

The neuron-specific RNA-binding protein ELAV regulates neuroglian alternative splicing in neurons and binds directly to its pre-mRNA
- PMID: 11581160
- PMCID: PMC312793
- DOI: 10.1101/gad.903101
The neuron-specific RNA-binding protein ELAV regulates neuroglian alternative splicing in neurons and binds directly to its pre-mRNA
Abstract
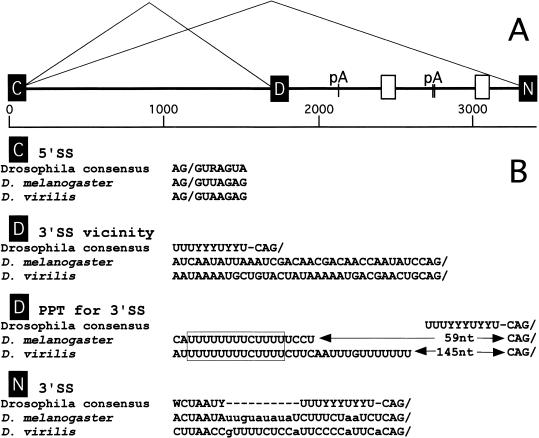
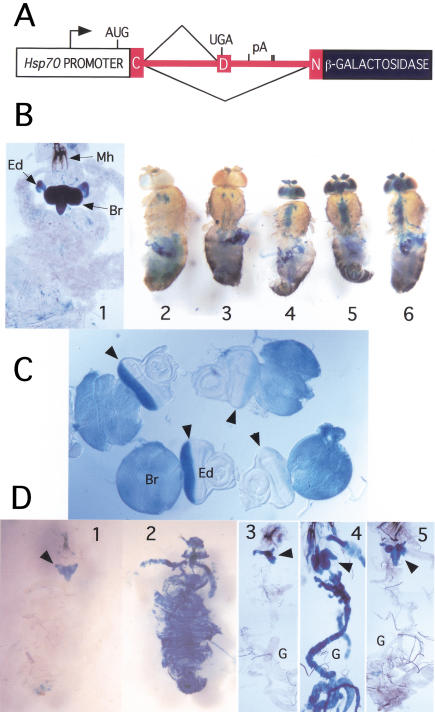
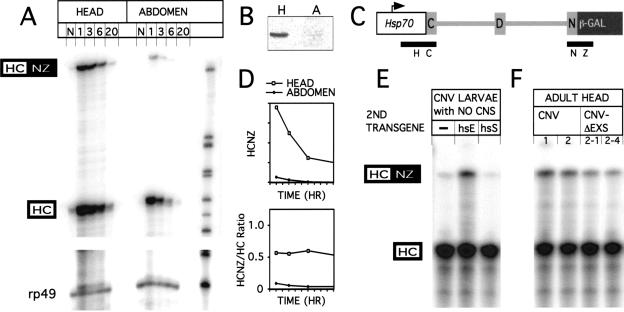
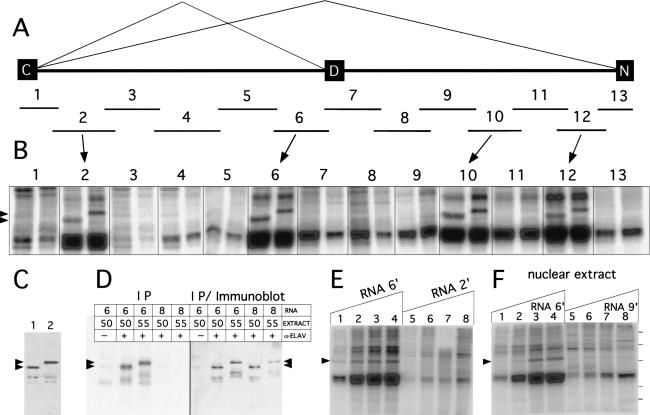
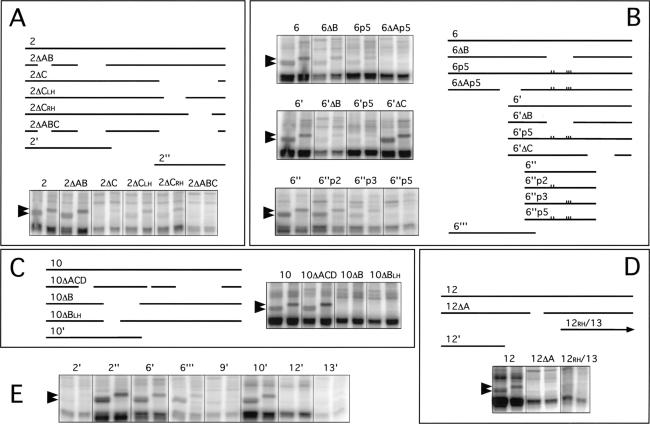
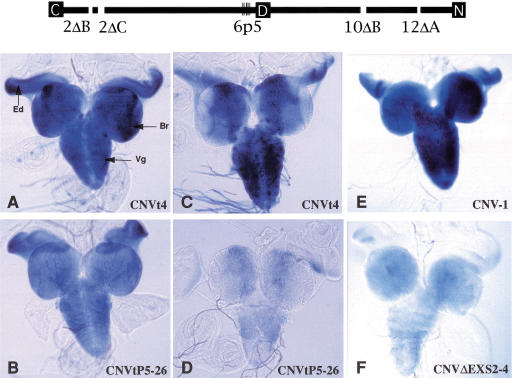
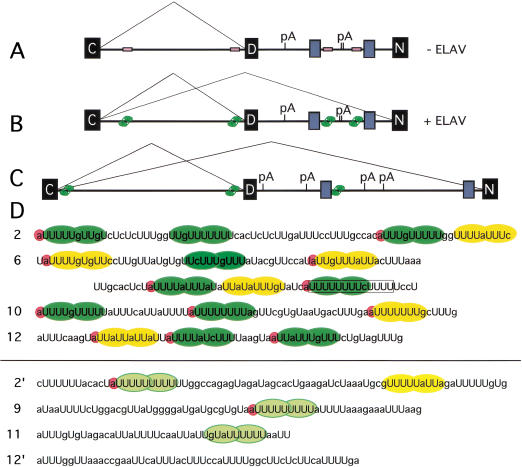
Drosophila melanogaster neural-specific protein, ELAV, has been shown to regulate the neural-specific splicing of three genes: neuroglian (nrg), erect wing, and armadillo. Alternative splicing of the nrg transcript involves alternative inclusion of a 3'-terminal exon. Here, using a minigene reporter, we show that the nrg alternatively spliced intron (nASI) has all the determinants required to recreate proper neural-specific RNA processing seen with the endogenous nrg transcript, including regulation by ELAV. An in vitro UV cross-linking assay revealed that ELAV from nuclear extracts cross-links to four distinct sites along the 3200 nucleotide long nASI; one EXS is positioned at the polypyrimidine tract of the default 3' splice site. ELAV cross-linking sites (EXSs) have in common long tracts of (U)-rich sequence rather than a precise consensus; moreover, each tract has at least two 8/10U elements; their importance is validated by mutant transgene reporter analysis. Further, we propose criteria for ELAV target sequence recognition based on the four EXSs, sites within the nASI that are (U) rich but do not cross-link with ELAV, and predicted EXSs from a phylogenetic comparison with Drosophila virilis nASI. These results suggest that ELAV regulates nrg alternative splicing by direct interaction with the nASI.
Figures
References
-
- Amara SG, Jonas V, Rosenfeld MG, Ong ES, Evans RM. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature. 1982;298:240–244. - PubMed
-
- Bashaw GJ, Baker BS. The regulation of the Drosophila msl-2 gene reveals a function for Sex-lethal in translational control. Cell. 1997;89:789–798. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases