

Regulation of inherently autoreactive VH4-34 B cells in the maintenance of human B cell tolerance
- PMID: 11581307
- PMCID: PMC200949
- DOI: 10.1172/JCI12462
Regulation of inherently autoreactive VH4-34 B cells in the maintenance of human B cell tolerance
Abstract
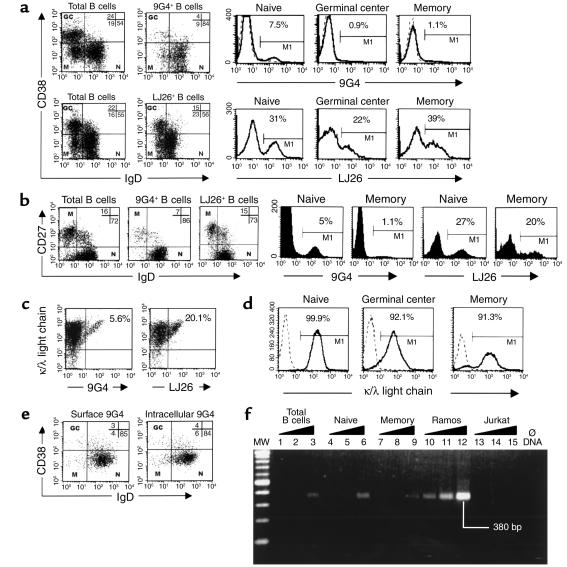
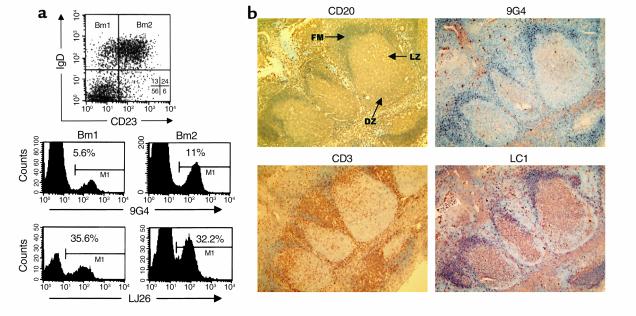
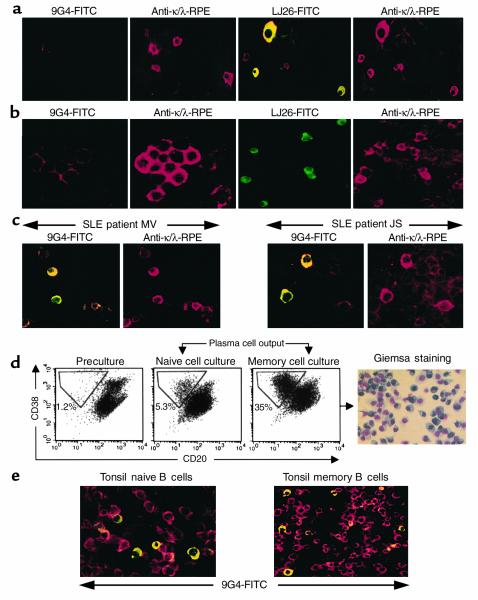
The study of human B cell tolerance has been hampered by difficulties in identifying a sizable population of autoreactive B lymphocytes whose fate could be readily determined. Hypothesizing that B cells expressing intrinsically autoreactive antibodies encoded by the VH4-34 heavy chain gene (VH4-34 cells) represent such a population, we tracked VH4-34 cells in healthy individuals. Here, we show that naive VH4-34 cells are positively selected and mostly restricted to the follicular mantle zone. Subsequently, these cells are largely excluded from the germinal centers and underrepresented in the memory compartment. In healthy donors but not in patients with systemic lupus erythematosus (SLE), these cells are prevented from differentiating into antibody-producing plasma cells. This blockade can be overcome ex vivo using cultures of naive and memory VH4-34 cells in the presence of CD70, IL-2, and IL-10. VH4-34 cells may therefore represent an experimentally useful surrogate for autoantibody transgenes and should prove valuable in studying human B cell tolerance in a physiological, polyclonal environment. Our initial results suggest that both positive and negative selection processes participate in the maintenance of tolerance in autoreactive human B cells at multiple checkpoints throughout B cell differentiation and that at least some censoring mechanisms are faulty in SLE.
Figures
Similar articles
-
Rituximab improves peripheral B cell abnormalities in human systemic lupus erythematosus.Arthritis Rheum. 2004 Nov;50(11):3580-90. doi: 10.1002/art.20592. Arthritis Rheum. 2004. PMID: 15529346 Clinical Trial.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
B cell subset composition segments clinically and serologically distinct groups in chronic cutaneous lupus erythematosus.Ann Rheum Dis. 2021 Sep;80(9):1190-1200. doi: 10.1136/annrheumdis-2021-220349. Epub 2021 Jun 3. Ann Rheum Dis. 2021. PMID: 34083207 Free PMC article.
Cited by
-
Fundamental roles of the innate-like repertoire of natural antibodies in immune homeostasis.Front Immunol. 2013 Feb 5;4:4. doi: 10.3389/fimmu.2013.00004. eCollection 2013. Front Immunol. 2013. PMID: 23386848 Free PMC article.
-
Loss of an IgG plasma cell checkpoint in patients with lupus.J Allergy Clin Immunol. 2019 Apr;143(4):1586-1597. doi: 10.1016/j.jaci.2018.10.041. Epub 2018 Nov 13. J Allergy Clin Immunol. 2019. PMID: 30439406 Free PMC article.
-
Primary cystic lung light chain deposition disease: a clinicopathologic entity derived from unmutated B cells with a stereotyped IGHV4-34/IGKV1 receptor.Blood. 2008 Sep 1;112(5):2004-12. doi: 10.1182/blood-2007-11-123596. Epub 2008 May 15. Blood. 2008. PMID: 18483396 Free PMC article.
-
CLL stereotyped B-cell receptor immunoglobulin sequences are recurrent in the B-cell repertoire of healthy individuals: Apparent lack of central and early peripheral tolerance censoring.Front Oncol. 2023 Mar 17;13:1112879. doi: 10.3389/fonc.2023.1112879. eCollection 2023. Front Oncol. 2023. PMID: 37007084 Free PMC article.
-
Autoantibody-mediated impairment of DNASE1L3 activity in sporadic systemic lupus erythematosus.J Exp Med. 2021 May 3;218(5):e20201138. doi: 10.1084/jem.20201138. J Exp Med. 2021. PMID: 33783474 Free PMC article.
References
-
- Burnet FM. The concept of immunological surveillance. Prog Exp Tumor Res. 1970;13:1–27. - PubMed
-
- Goodnow CC, Adelstein S, Basten A. The need for central and peripheral tolerance in the B cell repertoire. Science. 1990;248:1373–1379. - PubMed
-
- Nemazee D, et al. Clonal deletion of autospecific B lymphocytes. Immunol Rev. 1991;122:117–132. - PubMed
-
- Erikson J, et al. Expression of anti-DNA immunoglobulin transgenes in non-autoimmune mice. Nature. 1991;349:331–334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
