

Aortic valve endothelial cells undergo transforming growth factor-beta-mediated and non-transforming growth factor-beta-mediated transdifferentiation in vitro
- PMID: 11583961
- PMCID: PMC1850524
- DOI: 10.1016/s0002-9440(10)62520-5
Aortic valve endothelial cells undergo transforming growth factor-beta-mediated and non-transforming growth factor-beta-mediated transdifferentiation in vitro
Abstract
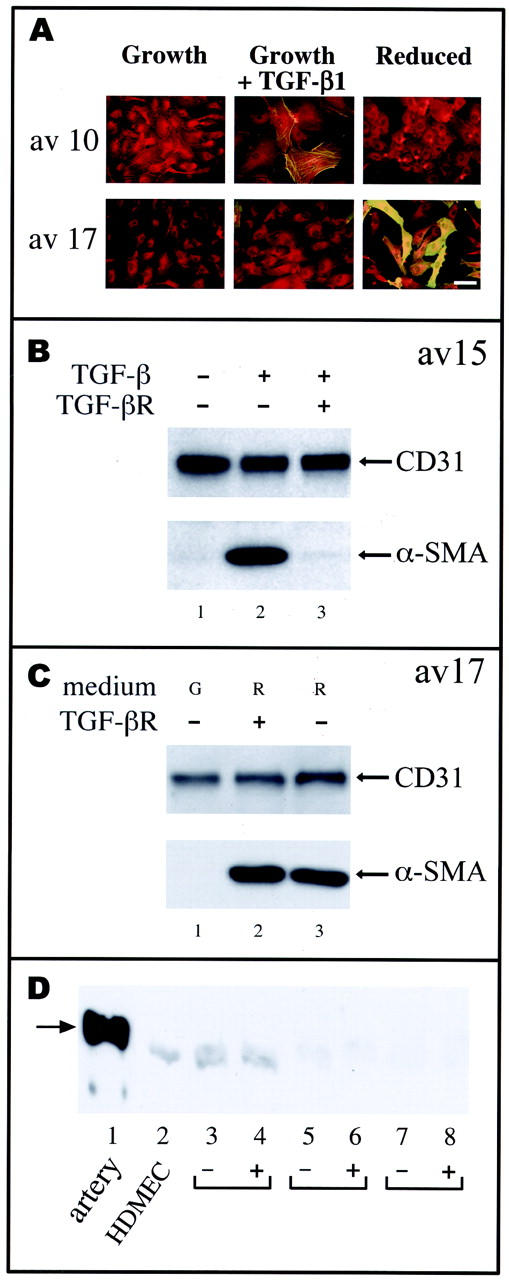
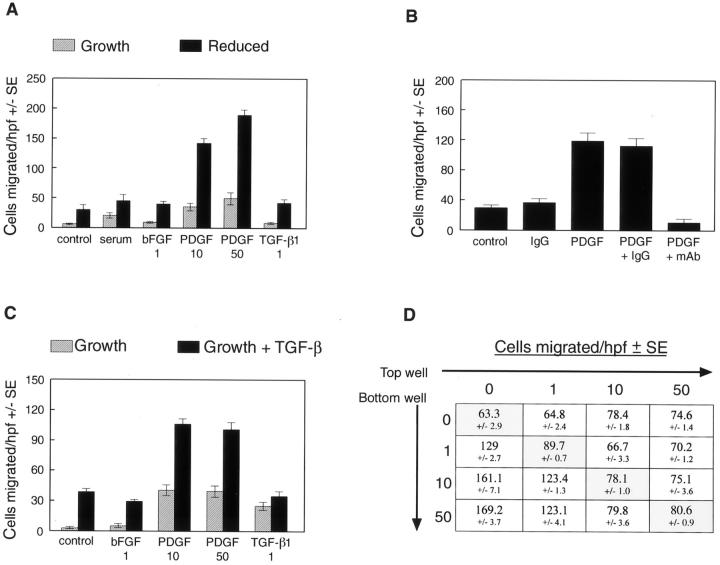
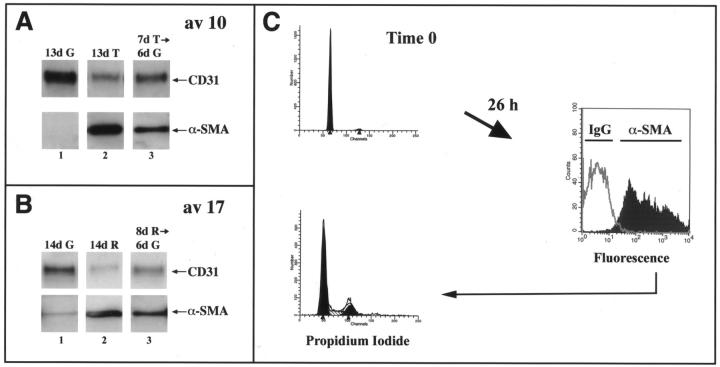
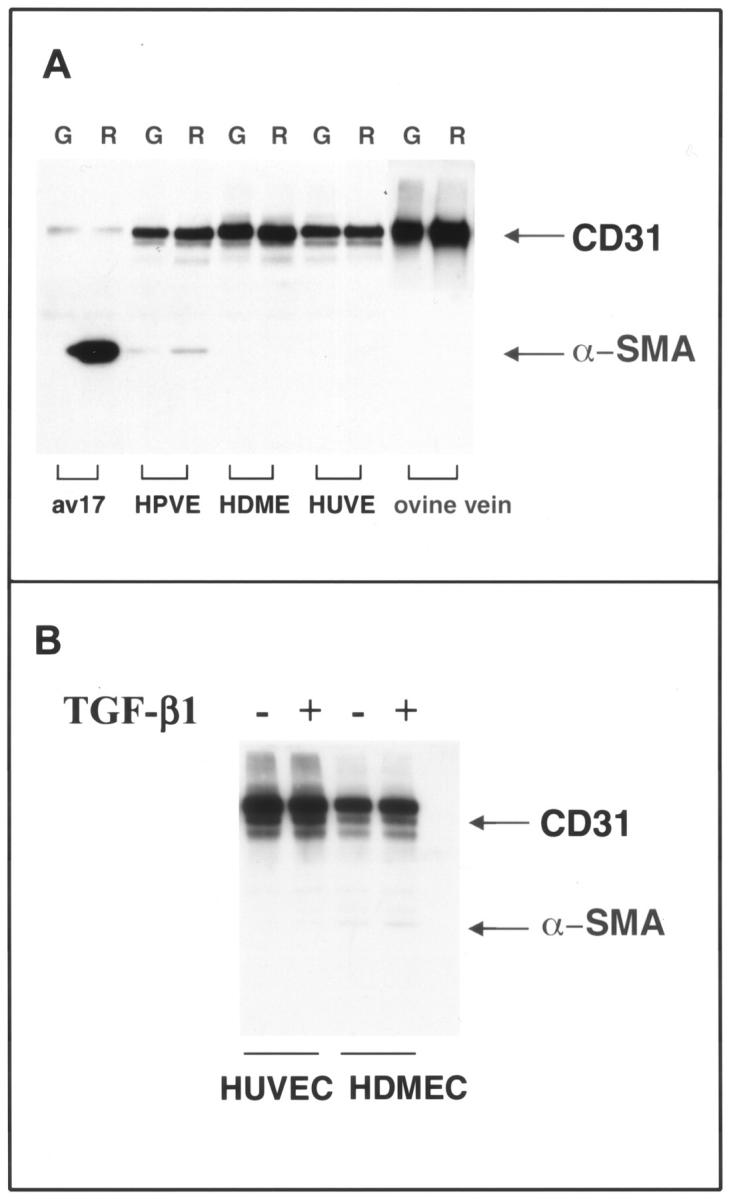
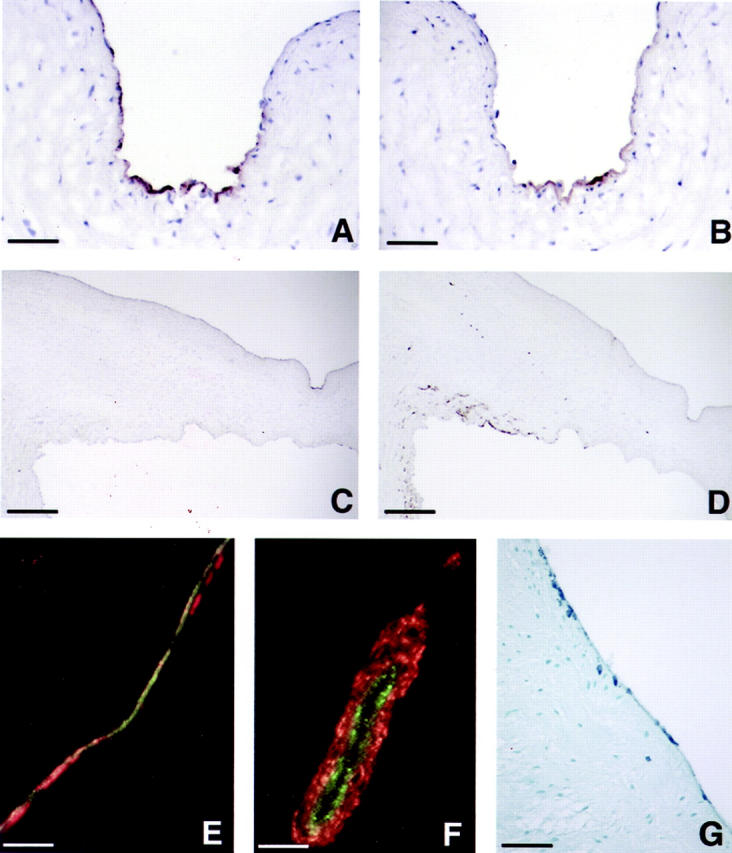
Cardiac valves arise from endocardial cushions, specialized regions of the developing heart that are formed by an endothelial-to-mesenchymal cell transdifferentiation. Whether and to what extent this transdifferentiation is retained in mature heart valves is unknown. Herein we show that endothelial cells from mature valves can transdifferentiate to a mesenchymal phenotype. Using induction of alpha-smooth muscle actin (alpha-SMA), an established marker for this process, two distinct pathways of transdifferentiation were identified in clonally derived endothelial cell populations isolated from ovine aortic valve leaflets. alpha-SMA expression was induced by culturing clonal endothelial cells in medium containing either transforming growth factor-beta or low levels of serum and no basic fibroblast growth factor. Cells induced to express alpha-SMA exhibited markedly increased migration in response to platelet-derived growth factor-BB, consistent with a mesenchymal phenotype. A population of the differentiated cells co-expressed CD31, an endothelial marker, along with alpha-SMA, as seen by double-label immunofluorescence. Similarly, this co-expression of endothelial markers and alpha-SMA was detected in a subpopulation of cells in frozen sections of aortic valves, suggesting the transdifferentiation may occur in vivo. Hence, the clonal populations of valvular endothelial cells described here provide a powerful in vitro model for dissecting molecular events that regulate valvular endothelium.
Figures

References
-
- Eisenberg LM, Markwald RR: Molecular regulation of atrioventricular valvuloseptal morphogenesis. Circ Res 1995, 77:1-6 - PubMed
-
- Schoen FJ, Levy RJ: Tissue heart valves: current challenges and future research perspectives. J Biomed Mat Res 1999, 47:439-465 - PubMed
-
- Mjaatvedt CH, Yamamura H, Wessels A, Ramsdell A, Turner D, Markwald RR: Mechanisms of segmentation, septation, and remodeling of the tubular heart: endocardial cushion fate and cardiac looping. Harvery RP Rosenthal N eds. Heart Development. 1999, :pp 159-177 Academic Press, San Diego
-
- Ramsdell AF, Markwald RR: Induction of endocardial cushion tissue in the avian heart is regulated in part by TGF-beta3-mediated autocrine signalling. Dev Biol 1997, 188:64-74 - PubMed
-
- Boyer AS, Ayerinskas II, Vincent EB, McKinney LA, Weeks DL, Runyan RB: TGF beta 2 and TGF beta 3 have separate and sequential activities during epithelial-mesenchymal cell transformation in the embryonic heart. Dev Biol 1999, 208:530-545 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
