

Side-specificity of olfactory learning in the honeybee: generalization between odors and sides
- PMID: 11584076
- PMCID: PMC311384
- DOI: 10.1101/lm.41401
Side-specificity of olfactory learning in the honeybee: generalization between odors and sides
Abstract
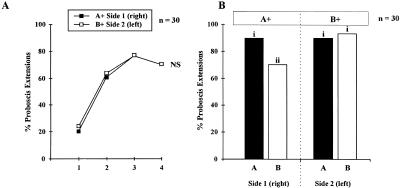
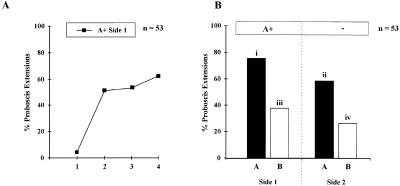
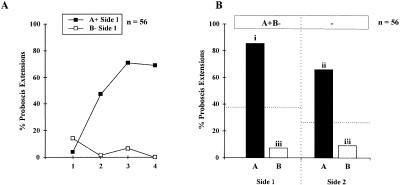
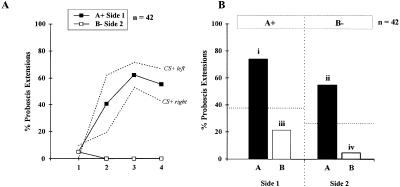
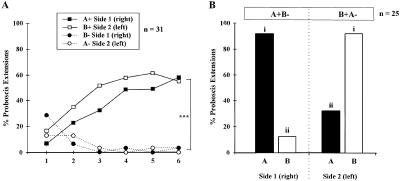
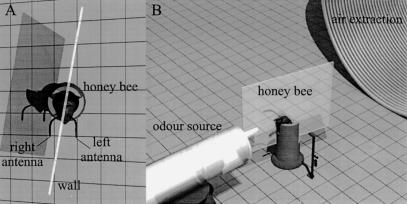
Honeybees (Apis mellifera) can be trained to associate an odor stimulus with a sucrose reward. The neural structures involved in the detection and integration of olfactory stimuli are represented bilaterally in the brain. Little is known about the respective roles of the two sides of the brain in olfactory learning. Does each side learn independently of the other, or do they communicate, and if so, to what extent and at what level of neural integration? We addressed these questions using the proboscis extension response (PER) conditioning paradigm applied in a preparation that allows the separation of the two input sides during olfactory stimulations. Bees conditioned to two odorants A and B, one being learned on each side (A+/B+ training), showed in extinction tests rather unspecific responses: They responded to both odorants on both sides. This could be attributable to either a transfer of the learned information between sides, or to a generalization between odorants on each side. By subjecting bees to conditioning on one side only (A+/0 training), we found that the learned information is indeed transferred between sides. However, when bees were trained explicitly to give opposite values to the two odorants on the two sides (A+B-/B+A- training), they showed clear side-specific response patterns to these odorants. These results are used in the elaboration of a functional model of laterality of olfactory learning and memory processing in the honeybee brain.
Figures
References
-
- Arnold G, Masson C, Budharugsa S. Comparative study of the antennal lobes and their afferent pathway in the worker bee and the drone (Apis mellifera) Cell Tissue Res. 1985;242:593–605.
-
- Chandra S, Smith BH. An analysis of synthetic processing of odor mixtures in the honeybee. J Exp Biol. 1998;201:3113–3121. - PubMed
-
- Erber J, Masuhr T, Menzel R. Localization of short-term memory in the brain of the bee, Apis mellifera. Physiol Entomol. 1980;5:343–358.
-
- Erber J, Pribbenow B, Grandy K, Kierzek S. Tactile motor learning in the antennal system of the honeybee (Apis mellifera L.) J Comp Physiol A. 1997;181:355–365.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources