

Molecular basis for telomere repeat divergence in budding yeast
- PMID: 11585910
- PMCID: PMC99902
- DOI: 10.1128/MCB.21.21.7277-7286.2001
Molecular basis for telomere repeat divergence in budding yeast
Abstract
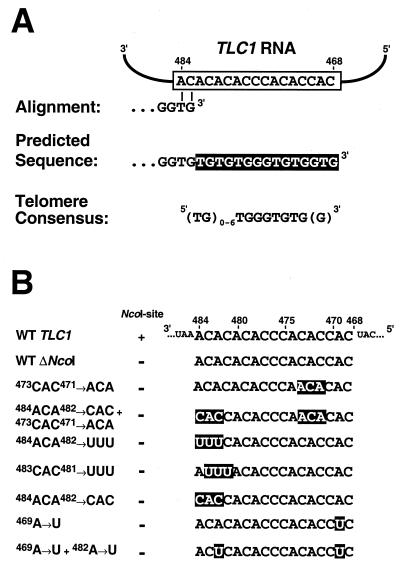
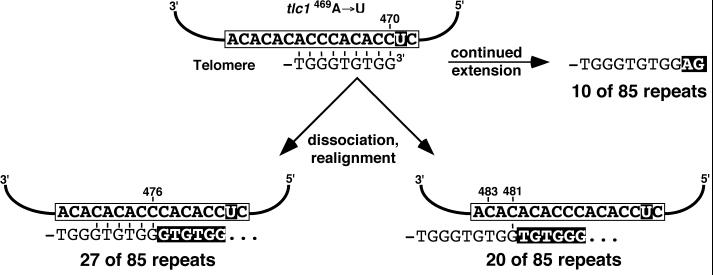
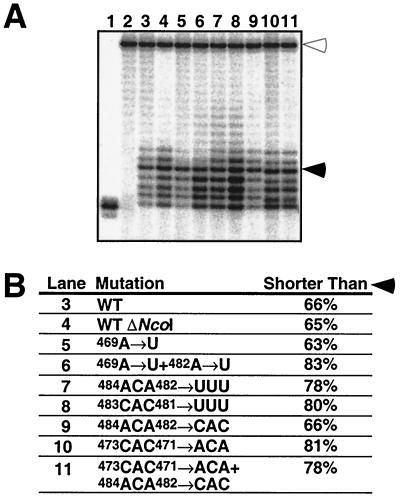
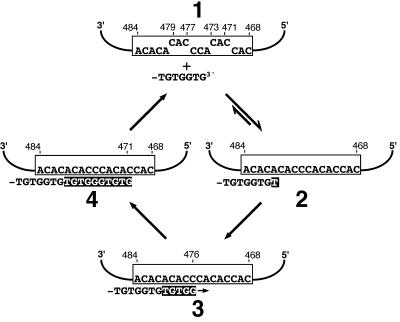
Telomerase is a ribonucleoprotein enzyme that adds repetitive sequences to the ends of linear chromosomes, thereby counteracting nucleotide loss due to incomplete replication. A short region of the telomerase RNA subunit serves as template for nucleotide addition onto the telomere 3' end. Although Saccharomyces cerevisiae contains only one telomerase RNA gene, telomere repeat sequences are degenerate in this organism. Based on a detailed analysis of the telomere sequences specified by wild-type and mutant RNA templates in vivo, we show that the divergence of telomere repeats is due to abortive reverse transcription in the 3' and 5' regions of the template and due to the alignment of telomeres in multiple registers within the RNA template. Through the interpretation of wild-type telomere sequences, we identify nucleotides in the template that are not accessible for base pairing during substrate annealing. Rather, these positions become available as templates for reverse transcription only after alignment with adjacent nucleotides has occurred, indicating that a conformational change takes place upon substrate binding. We also infer that the central part of the template region is reverse transcribed processively. The inaccessibility of certain template positions for alignment and the processive polymerization of the central template portion may serve to reduce the possible repeat diversification and enhance the incorporation of binding sites for Rap1p, the telomere binding protein of budding yeast.
Figures

Similar articles
-
The telomere and telomerase: nucleic acid-protein complexes acting in a telomere homeostasis system. A review.Biochemistry (Mosc). 1997 Nov;62(11):1196-201. Biochemistry (Mosc). 1997. PMID: 9467842 Review.
-
Telomerase is an unusual RNA-containing enzyme. A review.Biochemistry (Mosc). 1997 Nov;62(11):1206-15. Biochemistry (Mosc). 1997. PMID: 9467844 Review.
-
Saccharomyces cerevisiae telomerase subunit Est3p binds DNA and RNA and stimulates unwinding of RNA/DNA heteroduplexes.FEBS Lett. 2006 Aug 21;580(19):4683-90. doi: 10.1016/j.febslet.2006.07.048. Epub 2006 Jul 24. FEBS Lett. 2006. PMID: 16884717
-
Large telomerase RNA, telomere length heterogeneity and escape from senescence in Candida glabrata.FEBS Lett. 2009 Nov 19;583(22):3605-10. doi: 10.1016/j.febslet.2009.10.034. Epub 2009 Oct 17. FEBS Lett. 2009. PMID: 19840797
-
Telomere maintenance without telomerase.Oncogene. 2002 Jan 21;21(4):522-31. doi: 10.1038/sj.onc.1205079. Oncogene. 2002. PMID: 11850777 Review.
Cited by
-
Progressive Rearrangement of Telomeric Sequences Added to Both the ITR Ends of the Yeast Linear pGKL Plasmid.Biol Proced Online. 2003;5:29-42. doi: 10.1251/bpo44. Epub 2003 Feb 17. Biol Proced Online. 2003. PMID: 12734558 Free PMC article.
-
The many facets of homologous recombination at telomeres.Microb Cell. 2015 Jul 30;2(9):308-321. doi: 10.15698/mic2015.09.224. Microb Cell. 2015. PMID: 28357308 Free PMC article. Review.
-
Telomerase can act as a template- and RNA-independent terminal transferase.Proc Natl Acad Sci U S A. 2005 Jul 12;102(28):9778-83. doi: 10.1073/pnas.0502252102. Epub 2005 Jun 30. Proc Natl Acad Sci U S A. 2005. PMID: 15994230 Free PMC article.
-
TeloSearchLR: an algorithm to detect novel telomere repeat motifs using long sequencing reads.G3 (Bethesda). 2025 Jun 4;15(6):jkaf062. doi: 10.1093/g3journal/jkaf062. G3 (Bethesda). 2025. PMID: 40169380 Free PMC article.
-
Tying up the Ends: Plasticity in the Recognition of Single-Stranded DNA at Telomeres.Biochemistry. 2016 Sep 27;55(38):5326-40. doi: 10.1021/acs.biochem.6b00496. Epub 2016 Sep 15. Biochemistry. 2016. PMID: 27575340 Free PMC article.
References
-
- Bertuch A, Lundblad V. Telomeres and double-strand breaks—trying to make ends meet. Trends Cell Biol. 1998;8:339–342. - PubMed
-
- Bryan T M, Goodrich K J, Cech T R. A mutant of Tetrahymena telomerase reverse transcriptase with increased processivity. J Biol Chem. 2000;275:24199–24207. - PubMed
-
- Cohn M, Blackburn E H. Telomerase in yeast. Science. 1995;269:396–400. - PubMed
-
- Cohn M, McEachern M J, Blackburn E H. Telomeric sequence diversity within the genus Saccharomyces. Curr Genet. 1998;33:83–91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases