

claudin-18, a novel downstream target gene for the T/EBP/NKX2.1 homeodomain transcription factor, encodes lung- and stomach-specific isoforms through alternative splicing
- PMID: 11585919
- PMCID: PMC99911
- DOI: 10.1128/MCB.21.21.7380-7390.2001
claudin-18, a novel downstream target gene for the T/EBP/NKX2.1 homeodomain transcription factor, encodes lung- and stomach-specific isoforms through alternative splicing
Abstract
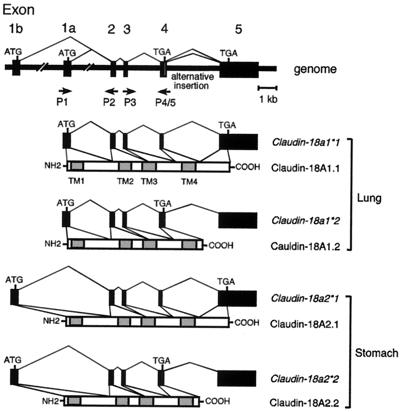
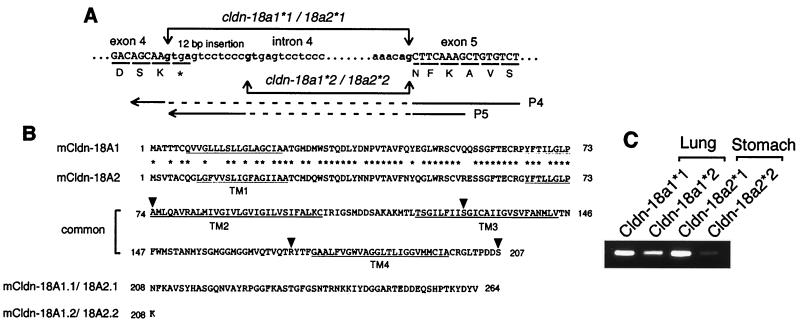
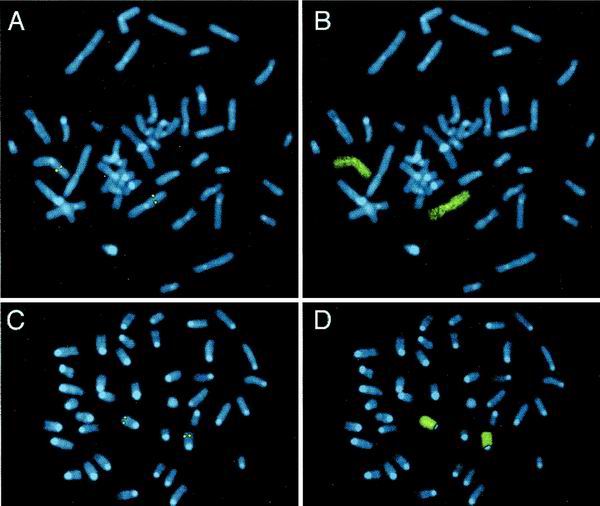
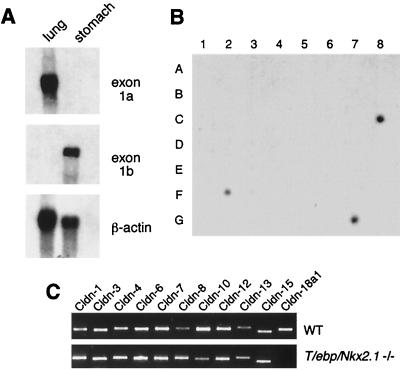
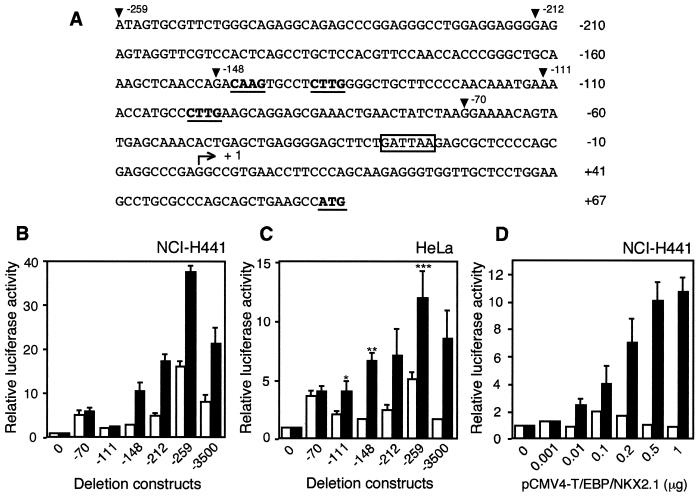
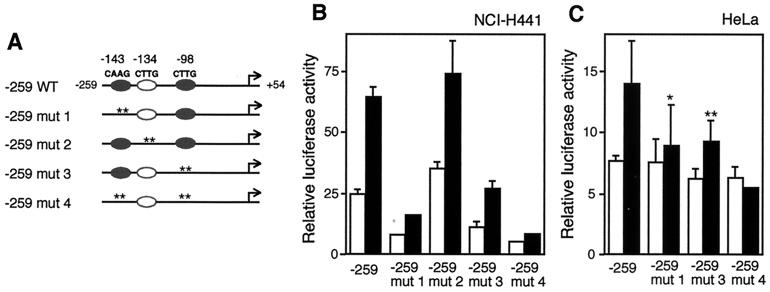
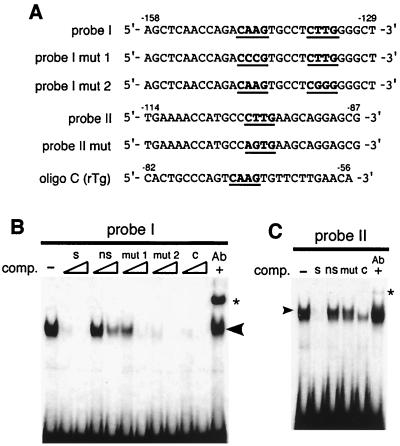
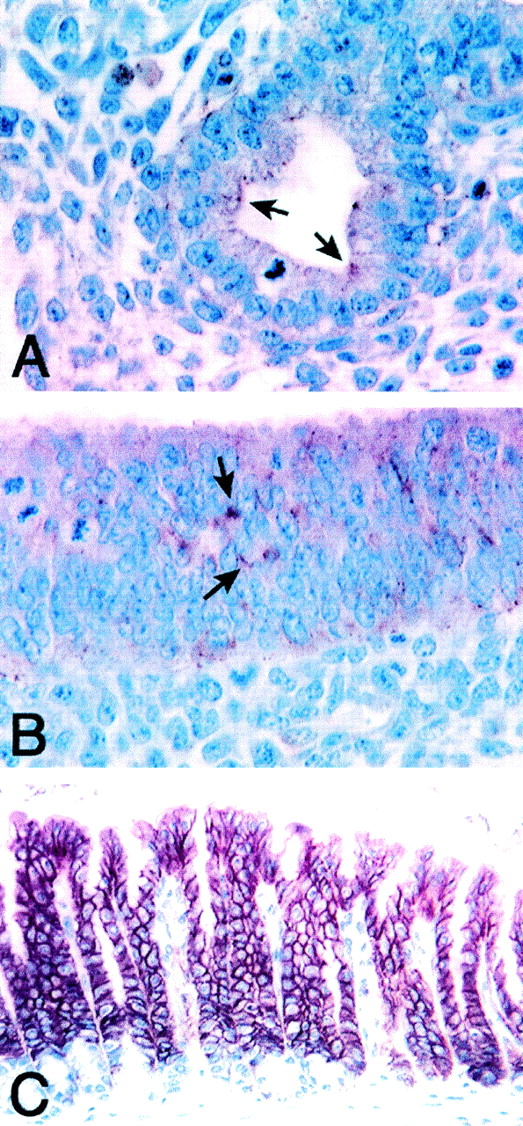
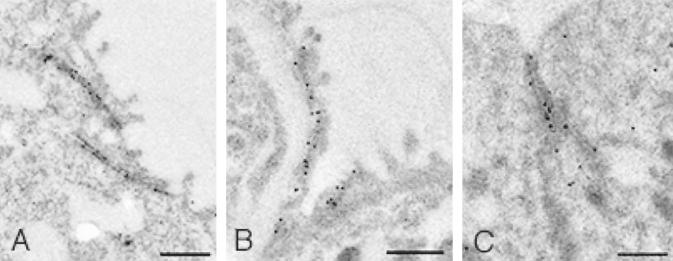
T/EBP/NKX2.1, a member of the NKX family of homeodomain-containing transcription factors, regulates the expression of a number of genes in lung and thyroid. Here we describe the isolation and characterization of a novel target gene, termed claudin-18, that is down-regulated in the lungs of T/ebp/Nkx2.1-null mouse embryos. The gene product exhibits an amino acid sequence similar to those of the claudin multigene family of proteins that constitute tight junction strands in epithelial cells. The gene was localized by fluorescence in situ hybridization to mouse chromosome 9 at region 9E3-F1 and to human chromosome 3 at region 3q21-23. The claudin-18 gene has two promoters, each with its own unique exon 1 that is spliced to common exons 2 through 5. Alternative usage of these promoters leads to production of lung and stomach-specific transcripts. The downstream lung-specific promoter contains two T/EBP/NKX2.1 binding sites responsible for trans activation of the gene by T/EBP/NKX2.1 in lung cells. Only claudin-18 was down-regulated in T/ebp/Nkx2.1-null embryo lungs among 11 claudin transcripts examined. Furthermore, the claudin-18 transcript has an alternative 12-bp insertion derived from the 5' end of intron 4, which produces a C-terminally truncated isoform in lung and stomach. Immunohistochemistry demonstrated complete membrane localization of claudin-18 with small focal dots in the lung and stomach epithelial cells. Immunogold electron microscopy analysis revealed that claudin-18 is concentrated at the cell-cell borders of epithelial cells. These unique features suggest a potentially important role for claudin-18 in the structure and function of tight junctions in lung and stomach.
Figures
References
-
- Bohinski R J, Di Lauro R, Whitsett J A. The lung-specific surfactant protein B gene promoter is a target for thyroid transcription factor 1 and hepatocyte nuclear factor 3, indicating common factors for organ-specific gene expression along the foregut axis. Mol Cell Biol. 1994;14:5671–5681. - PMC - PubMed
-
- Bruno M D, Bohinski R J, Huelsman K M, Whitsett J A, Korfhagen T R. Lung cell-specific expression of the murine surfactant protein A (SP-A) gene is mediated by interactions between the SP-A promoter and thyroid transcription factor-1. J Biol Chem. 1995;270:6531–6536. - PubMed
-
- Cardoso W V. Lung morphogenesis revisited: old facts, current ideas. Dev Dyn. 2000;219:121–130. - PubMed
-
- Cereijido M, Shoshani L, Contreras R G. Molecular physiology and pathophysiology of tight junctions. I. Biogenesis of tight junctions and epithelial polarity. Am J Physiol Gastrointest Liver Physiol. 2000;279:G477–G482. - PubMed
-
- Cereijido M, Valdes J, Shoshani L, Contreras R G. Role of tight junctions in establishing and maintaining cell polarity. Annu Rev Physiol. 1998;60:161–177. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases