

Downregulation of the Ras-mitogen-activated protein kinase pathway by the EphB2 receptor tyrosine kinase is required for ephrin-induced neurite retraction
- PMID: 11585923
- PMCID: PMC99915
- DOI: 10.1128/MCB.21.21.7429-7441.2001
Downregulation of the Ras-mitogen-activated protein kinase pathway by the EphB2 receptor tyrosine kinase is required for ephrin-induced neurite retraction
Abstract
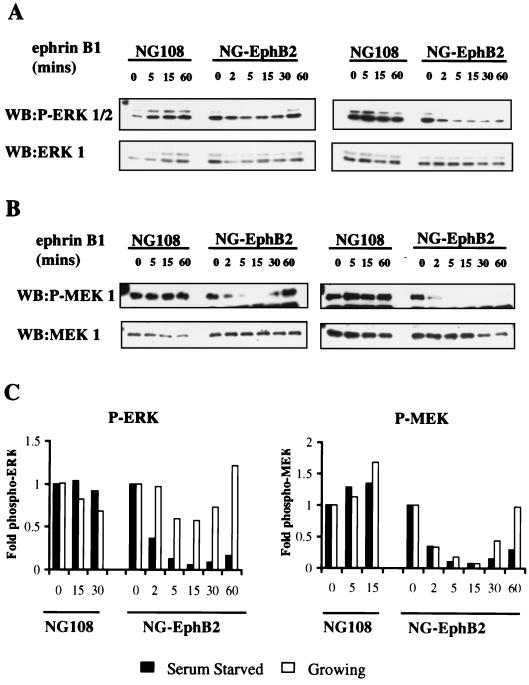
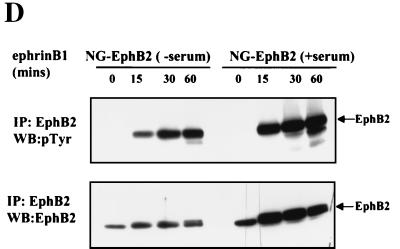
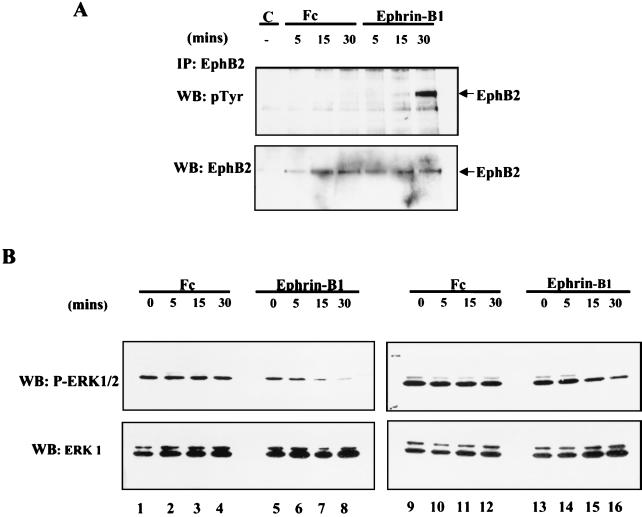
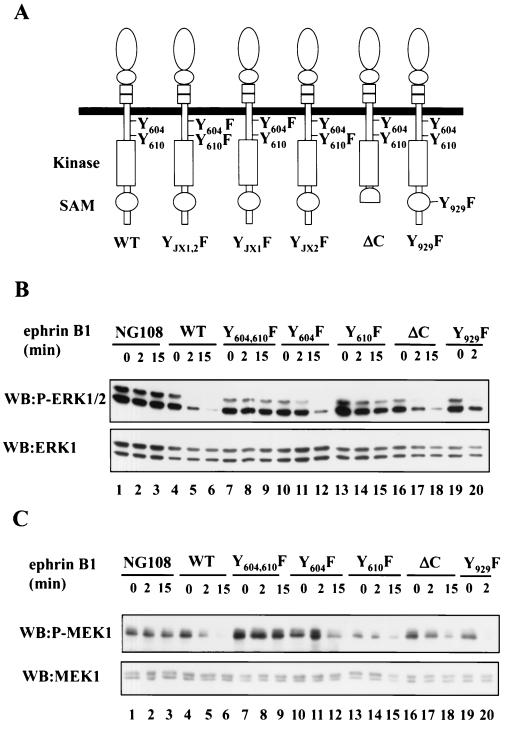
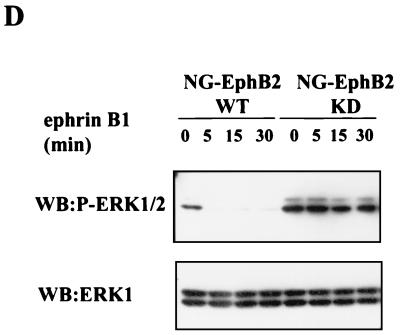
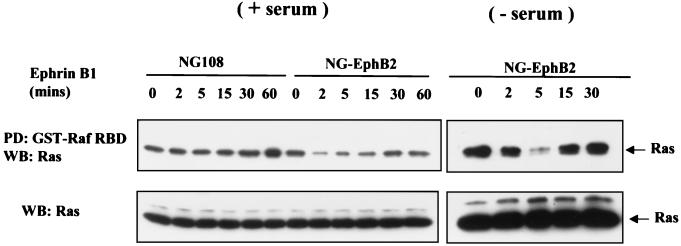
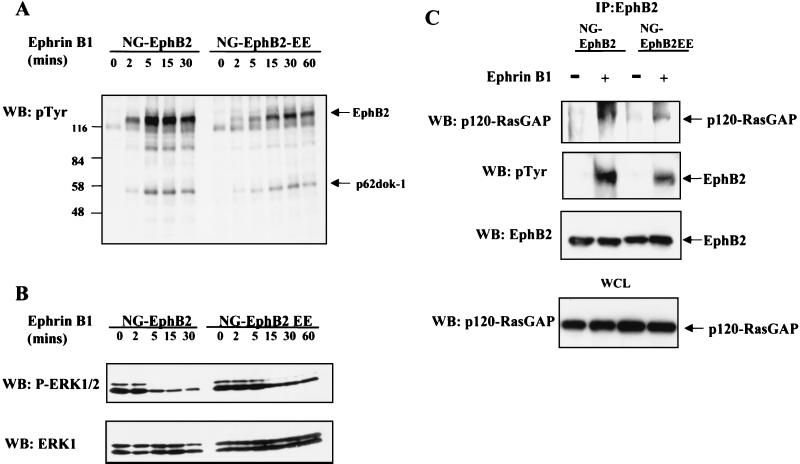
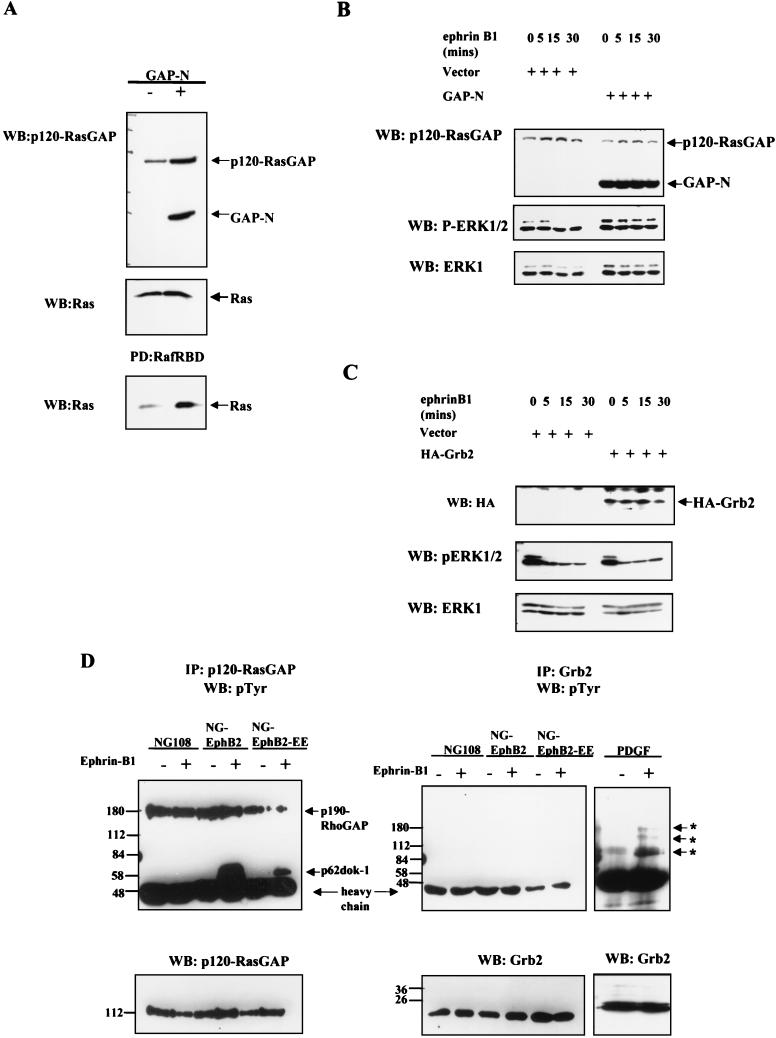
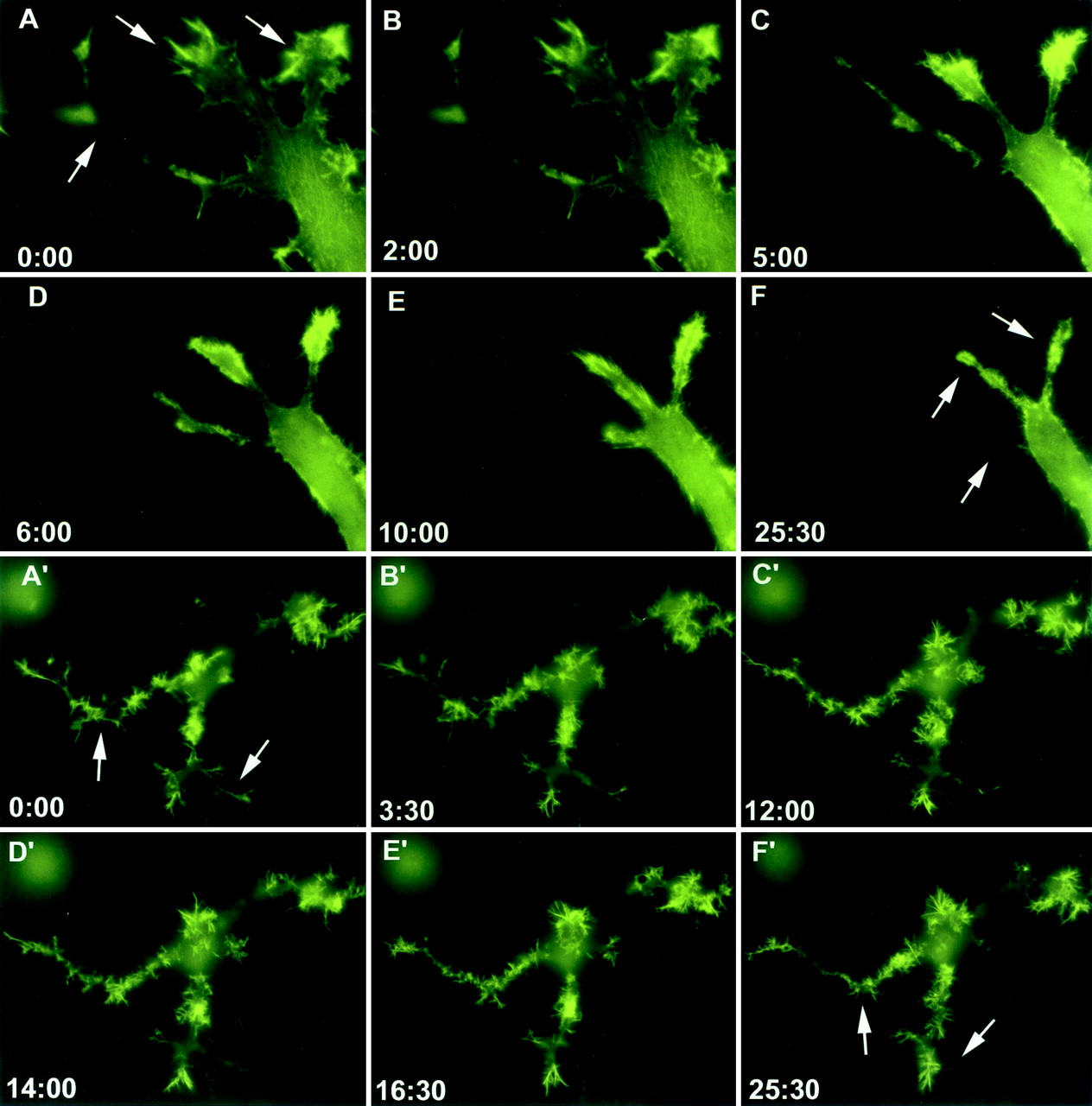
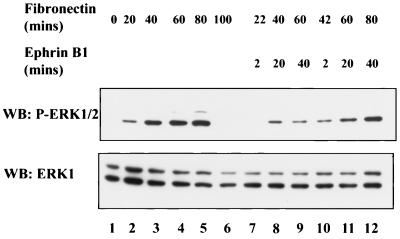
Activation of the EphB2 receptor tyrosine kinase by clustered ephrin-B1 induces growth cone collapse and neurite retraction in differentiated NG108 neuronal cells. We have investigated the cytoplasmic signaling events associated with EphB2-induced cytoskeletal reorganization in these neuronal cells. We find that unlike other receptor tyrosine kinases, EphB2 induces a pronounced downregulation of GTP-bound Ras and consequently of the extracellular signal-regulated kinase (ERK) mitogen-activated protein kinase (MAPK) pathway. A similar inhibition of the Ras-MAPK pathway was observed on stimulation of endogenous EphB2 in COS-1 cells. Inactivation of Ras, induced by ephrin B1 stimulation of NG108 neuronal cells, requires EphB2 tyrosine kinase activity and is blocked by a truncated form of p120-Ras GTPase-activating protein (p120-RasGAP), suggesting that EphB2 signals through the SH2 domain protein p120-RasGAP to inhibit the Ras-MAPK pathway. Suppression of Ras activity appears functionally important, since expression of a constitutively active variant of Ras impaired the ability of EphB2 to induce neurite retraction. In addition, EphB2 attenuated the elevation in ERK activation induced by attachment of NG108 cells to fibronectin, indicating that the EphB2 receptor can modulate integrin signaling to the Ras GTPase. These results suggest that a primary function of EphB2, a member of the most populous family of receptor tyrosine kinases, is to inactivate the Ras-MAPK pathway in a fashion that contributes to cytoskeletal reorganization and adhesion responses in neuronal growth cones.
Figures
Similar articles
-
Juxtamembrane tyrosine residues couple the Eph family receptor EphB2/Nuk to specific SH2 domain proteins in neuronal cells.EMBO J. 1997 Jul 1;16(13):3877-88. doi: 10.1093/emboj/16.13.3877. EMBO J. 1997. PMID: 9233798 Free PMC article.
-
Activation of EphA receptor tyrosine kinase inhibits the Ras/MAPK pathway.Nat Cell Biol. 2001 May;3(5):527-30. doi: 10.1038/35074604. Nat Cell Biol. 2001. PMID: 11331884
-
EphB ligand, ephrinB2, suppresses the VEGF- and angiopoietin 1-induced Ras/mitogen-activated protein kinase pathway in venous endothelial cells.FASEB J. 2002 Jul;16(9):1126-8. doi: 10.1096/fj.01-0805fje. Epub 2002 May 21. FASEB J. 2002. PMID: 12039842
-
Protein tyrosine kinase-mediated pathways in G protein-coupled receptor signaling.Cell Biochem Biophys. 1999;30(3):369-87. doi: 10.1007/BF02738120. Cell Biochem Biophys. 1999. PMID: 10403057 Review.
-
Integrating the MAP kinase signal into the G1 phase cell cycle machinery.Bioessays. 2000 Sep;22(9):818-26. doi: 10.1002/1521-1878(200009)22:9<818::AID-BIES7>3.0.CO;2-6. Bioessays. 2000. PMID: 10944584 Review.
Cited by
-
Rewiring of PDZ domain-ligand interaction network contributed to eukaryotic evolution.PLoS Genet. 2012 Feb;8(2):e1002510. doi: 10.1371/journal.pgen.1002510. Epub 2012 Feb 9. PLoS Genet. 2012. PMID: 22346764 Free PMC article.
-
Quantitative phosphotyrosine proteomics of EphB2 signaling by stable isotope labeling with amino acids in cell culture (SILAC).J Proteome Res. 2006 Mar;5(3):581-8. doi: 10.1021/pr050362b. J Proteome Res. 2006. PMID: 16512673 Free PMC article.
-
Inactivation of Ras by p120GAP via focal adhesion kinase dephosphorylation mediates RGMa-induced growth cone collapse.J Neurosci. 2009 May 20;29(20):6649-62. doi: 10.1523/JNEUROSCI.0927-09.2009. J Neurosci. 2009. PMID: 19458235 Free PMC article.
-
Optogenetic activation of EphB2 receptor in dendrites induced actin polymerization by activating Arg kinase.Biol Open. 2017 Dec 15;6(12):1820-1830. doi: 10.1242/bio.029900. Biol Open. 2017. PMID: 29158322 Free PMC article.
-
Eph/ephrin signaling in cell-cell and cell-substrate adhesion.Front Biosci (Landmark Ed). 2012 Jan 1;17(2):473-97. doi: 10.2741/3939. Front Biosci (Landmark Ed). 2012. PMID: 22201756 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous