

The HOX homeodomain proteins block CBP histone acetyltransferase activity
- PMID: 11585930
- PMCID: PMC99922
- DOI: 10.1128/MCB.21.21.7509-7522.2001
The HOX homeodomain proteins block CBP histone acetyltransferase activity
Abstract
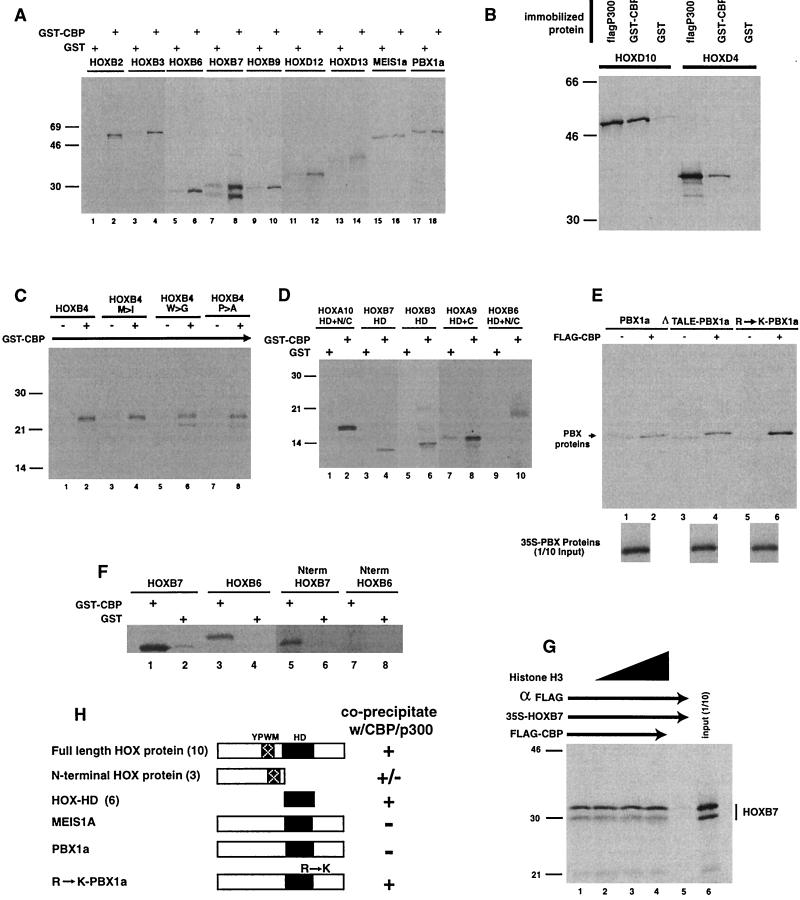
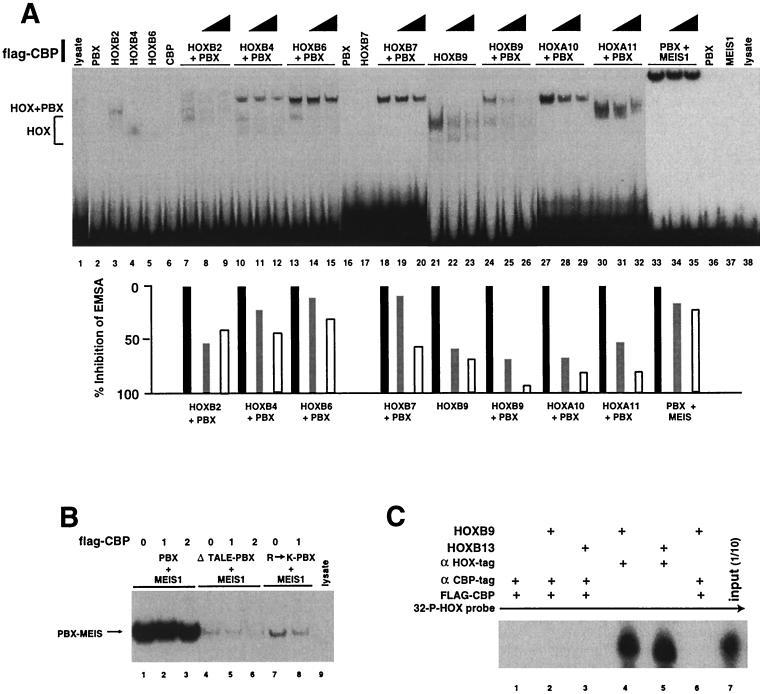
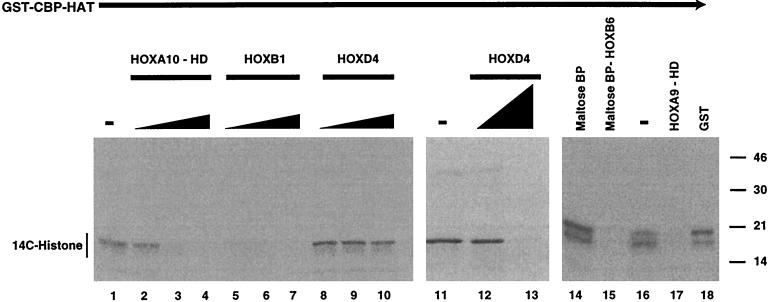
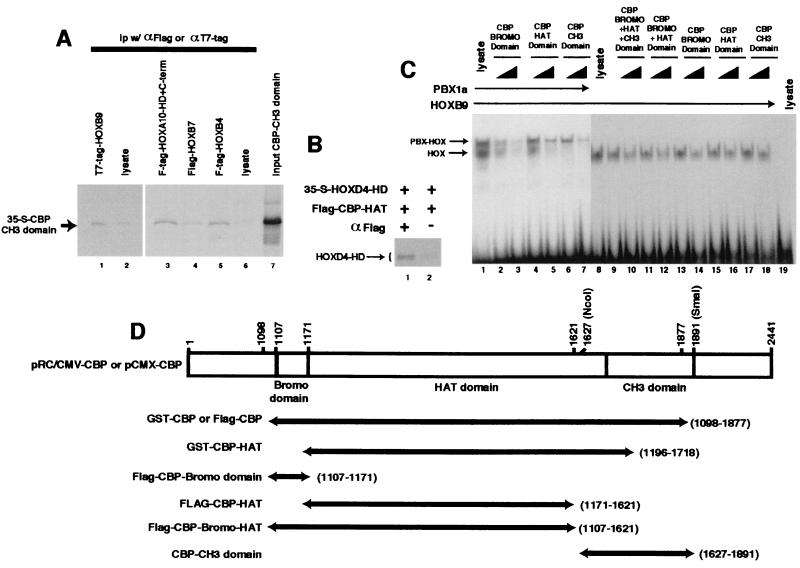
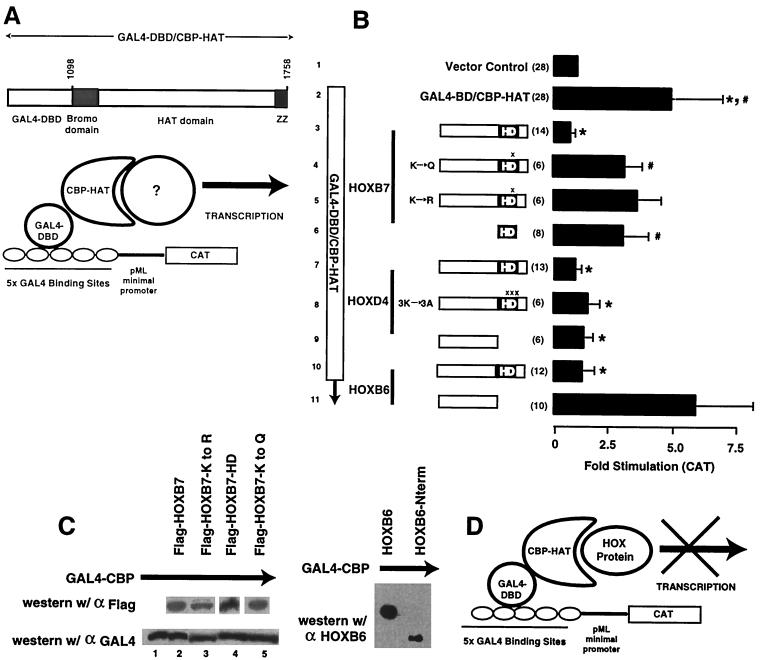
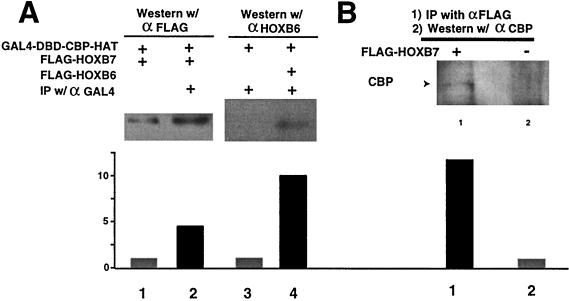
Despite the identification of PBC proteins as cofactors that provide DNA affinity and binding specificity for the HOX homeodomain proteins, HOX proteins do not demonstrate robust activity in transient-transcription assays and few authentic downstream targets have been identified for these putative transcription factors. During a search for additional cofactors, we established that each of the 14 HOX proteins tested, from 11 separate paralog groups, binds to CBP or p300. All six isolated homeodomain fragments tested bind to CBP, suggesting that the homeodomain is a common site of interaction. Surprisingly, CBP-p300 does not form DNA binding complexes with the HOX proteins but instead prevents their binding to DNA. The HOX proteins are not substrates for CBP histone acetyltransferase (HAT) but instead inhibit the activity of CBP in both in vitro and in vivo systems. These mutually inhibitory interactions are reflected by the inability of CBP to potentiate the low levels of gene activation induced by HOX proteins in a range of reporter assays. We propose two models for HOX protein function: (i) HOX proteins may function without CBP HAT to regulate transcription as cooperative DNA binding molecules with PBX, MEIS, or other cofactors, and (ii) the HOX proteins may inhibit CBP HAT activity and thus function as repressors of gene transcription.
Figures
Similar articles
-
Interaction and functional cooperation between the LIM protein FHL2, CBP/p300, and beta-catenin.Mol Cell Biol. 2004 Dec;24(24):10689-702. doi: 10.1128/MCB.24.24.10689-10702.2004. Mol Cell Biol. 2004. PMID: 15572674 Free PMC article.
-
MEIS C termini harbor transcriptional activation domains that respond to cell signaling.J Biol Chem. 2005 Mar 18;280(11):10119-27. doi: 10.1074/jbc.M413963200. Epub 2005 Jan 15. J Biol Chem. 2005. PMID: 15654074
-
HOXB6 protein is bound to CREB-binding protein and represses globin expression in a DNA binding-dependent, PBX interaction-independent process.J Biol Chem. 2004 Sep 17;279(38):39895-904. doi: 10.1074/jbc.M404132200. Epub 2004 Jul 21. J Biol Chem. 2004. PMID: 15269212
-
Hox cofactors in vertebrate development.Dev Biol. 2006 Mar 15;291(2):193-206. doi: 10.1016/j.ydbio.2005.10.032. Epub 2006 Mar 3. Dev Biol. 2006. PMID: 16515781 Review.
-
Hox transcription factors and their elusive mammalian gene targets.Heredity (Edinb). 2006 Aug;97(2):88-96. doi: 10.1038/sj.hdy.6800847. Epub 2006 May 24. Heredity (Edinb). 2006. PMID: 16721389 Review.
Cited by
-
Conditional knockout mice reveal distinct functions for the global transcriptional coactivators CBP and p300 in T-cell development.Mol Cell Biol. 2006 Feb;26(3):789-809. doi: 10.1128/MCB.26.3.789-809.2006. Mol Cell Biol. 2006. PMID: 16428436 Free PMC article.
-
TLX1/HOX11-induced hematopoietic differentiation blockade.Oncogene. 2007 Jun 14;26(28):4115-23. doi: 10.1038/sj.onc.1210185. Epub 2007 Jan 8. Oncogene. 2007. PMID: 17213805 Free PMC article.
-
Hox-dependent coordination of mouse cardiac progenitor cell patterning and differentiation.Elife. 2020 Aug 17;9:e55124. doi: 10.7554/eLife.55124. Elife. 2020. PMID: 32804075 Free PMC article.
-
Regulating histone acetyltransferases and deacetylases.EMBO Rep. 2003 Oct;4(10):944-7. doi: 10.1038/sj.embor.embor941. EMBO Rep. 2003. PMID: 14528264 Free PMC article. Review.
-
Construction of a HOXA11-AS-Interact Ed Network in Keloid Fibroblasts Using Integrated Bioinformatic Analysis and in Vitro Validation.Front Genet. 2022 Mar 31;13:844198. doi: 10.3389/fgene.2022.844198. eCollection 2022. Front Genet. 2022. PMID: 35432479 Free PMC article.
References
-
- Boyes J, Byfield P, Nakatani Y, Ogryzko V. Regulation of activity of the transcription factor GATA-1 by acetylation. Nature. 1998;396:594–598. - PubMed
-
- Brownell J E, Allis C D. Special HATs for special occasions: linking histone acetylation to chromatin assembly and gene activation. Curr Opin Genet Dev. 1996;6:176–184. - PubMed
-
- Burglin T. A comprehensive classification of homeobox genes. In: Duboule D, editor. Guidebook to the homeobox genes. Oxford, England: Oxford University Press; 1994. pp. 25–71.
-
- Chakravarti D, Ogryzko V, Kao H Y, Nash A, Chen H, Nakatani Y, Evans R M. A viral mechanism for inhibition of p300 and PCAF acetyltransferase activity. Cell. 1999;96:393–403. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous