

A model for gene therapy of human hereditary lymphedema
- PMID: 11592985
- PMCID: PMC60113
- DOI: 10.1073/pnas.221449198
A model for gene therapy of human hereditary lymphedema
Abstract
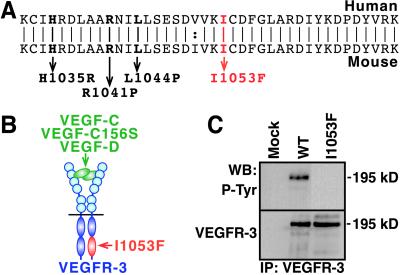
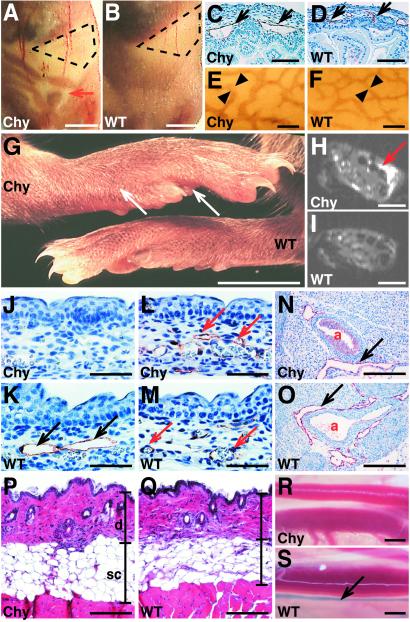
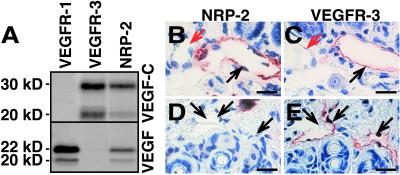
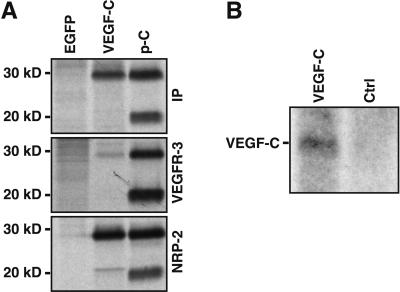
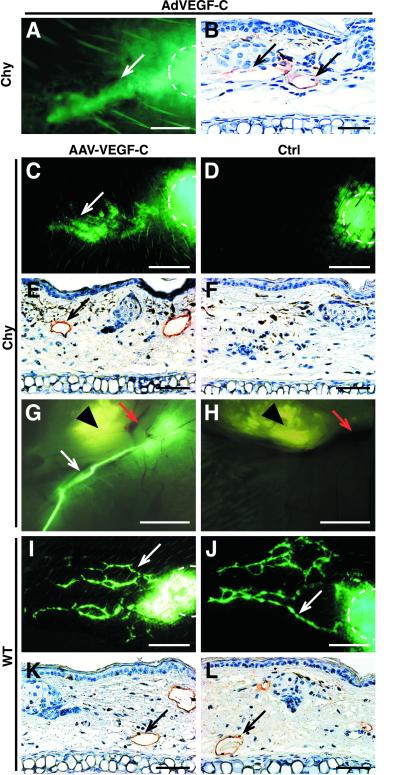
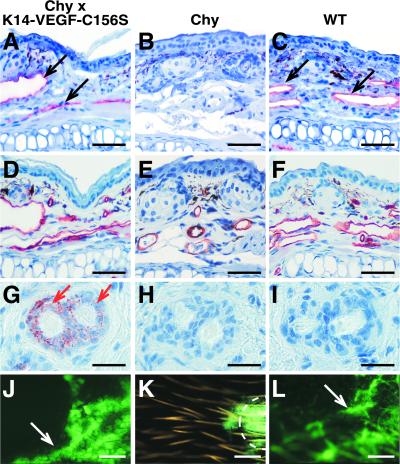
Primary human lymphedema (Milroy's disease), characterized by a chronic and disfiguring swelling of the extremities, is associated with heterozygous inactivating missense mutations of the gene encoding vascular endothelial growth factor C/D receptor (VEGFR-3). Here, we describe a mouse model and a possible treatment for primary lymphedema. Like the human patients, the lymphedema (Chy) mice have an inactivating Vegfr3 mutation in their germ line, and swelling of the limbs because of hypoplastic cutaneous, but not visceral, lymphatic vessels. Neuropilin (NRP)-2 bound VEGF-C and was expressed in the visceral, but not in the cutaneous, lymphatic endothelia, suggesting that it may participate in the pathogenesis of lymphedema. By using virus-mediated VEGF-C gene therapy, we were able to generate functional lymphatic vessels in the lymphedema mice. Our results suggest that growth factor gene therapy is applicable to human lymphedema and provide a paradigm for other diseases associated with mutant receptors.
Figures
References
-
- Witte M H, Way D L, Witte C L, Bernas M. In: Regulation of Angiogenesis. Goldberg I D, Rosen E M, editors. Basel: Birkhäuser; 1997. pp. 65–112.
-
- Ferrell R E, Levinson K L, Esman J H, Kimak M A, Lawrence E C, Barmada M M, Finegold D N. Hum Mol Genet. 1998;7:2073–2078. - PubMed
-
- Witte M H, Erickson R, Bernas M, Andrade M, Reiser F, Conlon W, Hoyme H E, Witte C L. Lymphology. 1998;31:145–155. - PubMed
-
- Karkkainen M J, Ferrell R E, Lawrence E C, Kimak M A, Levinson K L, McTigue M A, Alitalo K, Finegold D N. Nat Genet. 2000;25:153–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
