

Crystal structure of the central region of bovine fibrinogen (E5 fragment) at 1.4-A resolution
- PMID: 11593005
- PMCID: PMC59751
- DOI: 10.1073/pnas.211439798
Crystal structure of the central region of bovine fibrinogen (E5 fragment) at 1.4-A resolution
Abstract
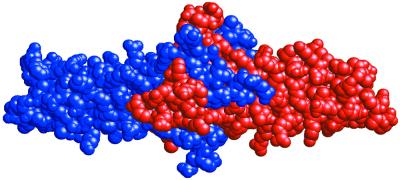
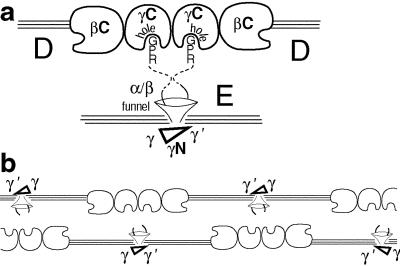
The high-resolution crystal structure of the N-terminal central region of bovine fibrinogen (a 35-kDa E(5) fragment) reveals a remarkable dimeric design. The two halves of the molecule bond together at the center in an extensive molecular "handshake" by using both disulfide linkages and noncovalent contacts. On one face of the fragment, the Aalpha and Bbeta chains from the two monomers form a funnel-shaped domain with an unusual hydrophobic cavity; here, on each of the two outer sides there appears to be a binding site for thrombin. On the opposite face, the N-terminal gamma chains fold into a separate domain. Despite the chemical identity of the two halves of fibrinogen, an unusual pair of adjacent disulfide bonds locally constrain the two gamma chains to adopt different conformations. The striking asymmetry of this domain may promote the known supercoiling of the protofibrils in fibrin. This information on the detailed topology of the E(5) fragment permits the construction of a more detailed model than previously possible for the critical trimolecular junction of the protofibril in fibrin.
Figures
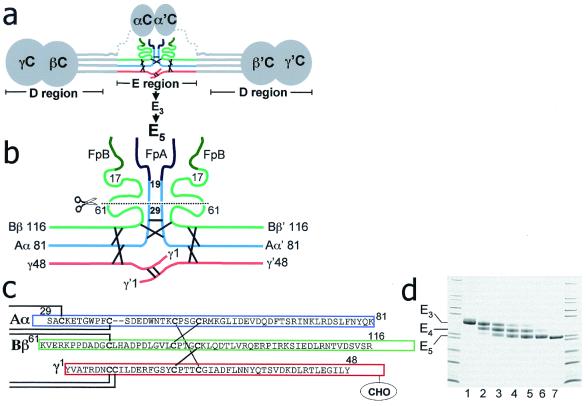
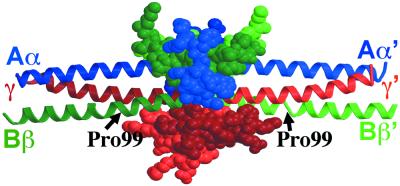
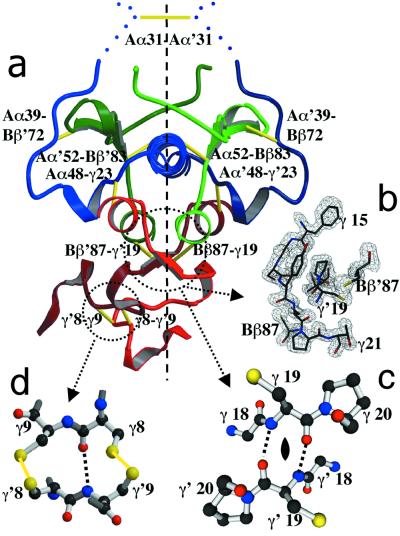
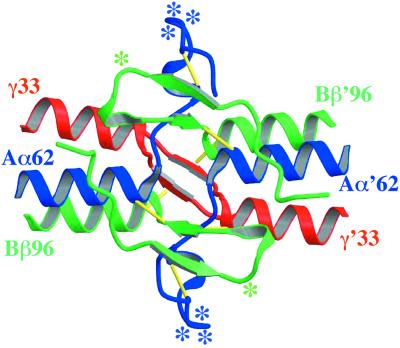
References
-
- Doolittle R F, Everse S J, Spraggon G. FASEB J. 1996;10:1464–1470. - PubMed
-
- Stryer L, Cohen C, Langridge R. Nature (London) 1963;197:793–794. - PubMed
-
- Weisel J W, Phillips G N, Jr, Cohen C. In: Molecular Biology of Fibrinogen and Fibrin. Mosesson M W, Doolittle R F, editors. New York: NY Acad. Sci.; 1983. pp. 367–379. - PubMed
-
- Hantgan R, McDonagh J, Hermans J. In: Molecular Biology of Fibrinogen and Fibrin. Mosesson M W, Doolittle R F, editors. New York: NY Acad. Sci.; 1983. pp. 344–366.
-
- Weisel J W, Veklich Y, Gorkun O. J Mol Biol. 1993;232:285–297. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
