

Plastid division is driven by a complex mechanism that involves differential transition of the bacterial and eukaryotic division rings
- PMID: 11595800
- PMCID: PMC139157
- DOI: 10.1105/tpc.010185
Plastid division is driven by a complex mechanism that involves differential transition of the bacterial and eukaryotic division rings
Abstract
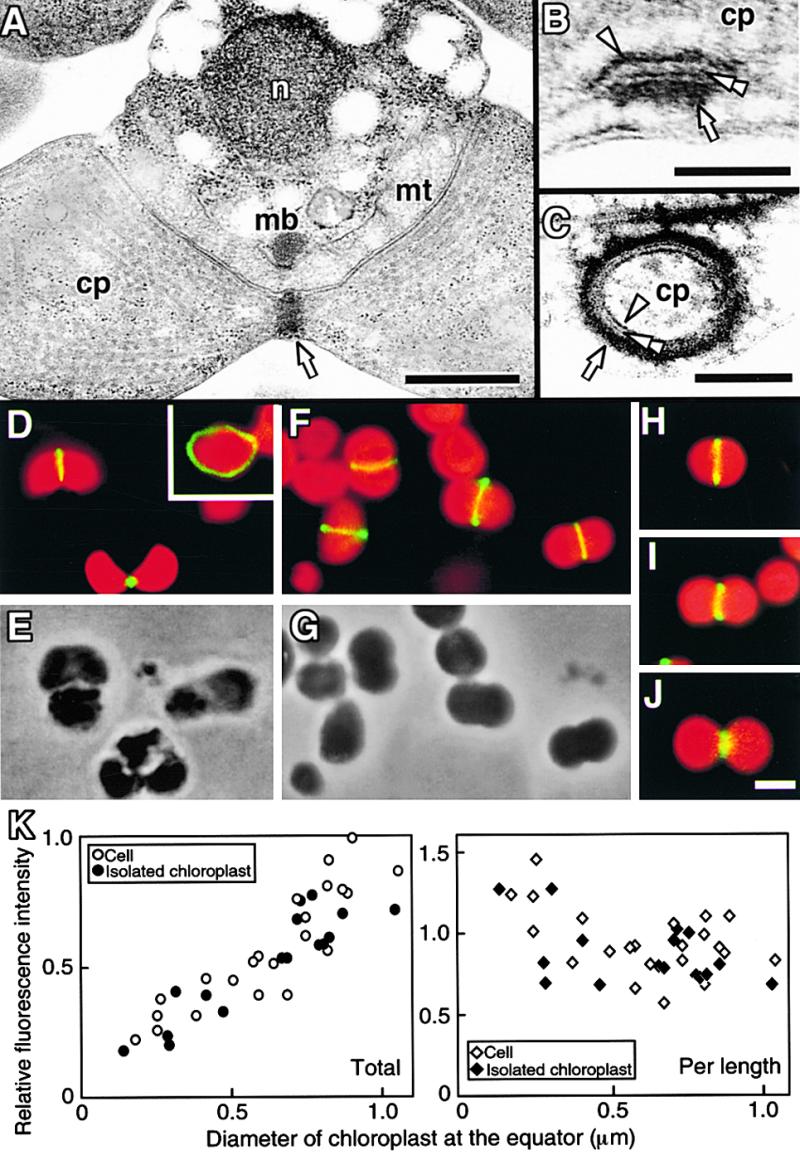
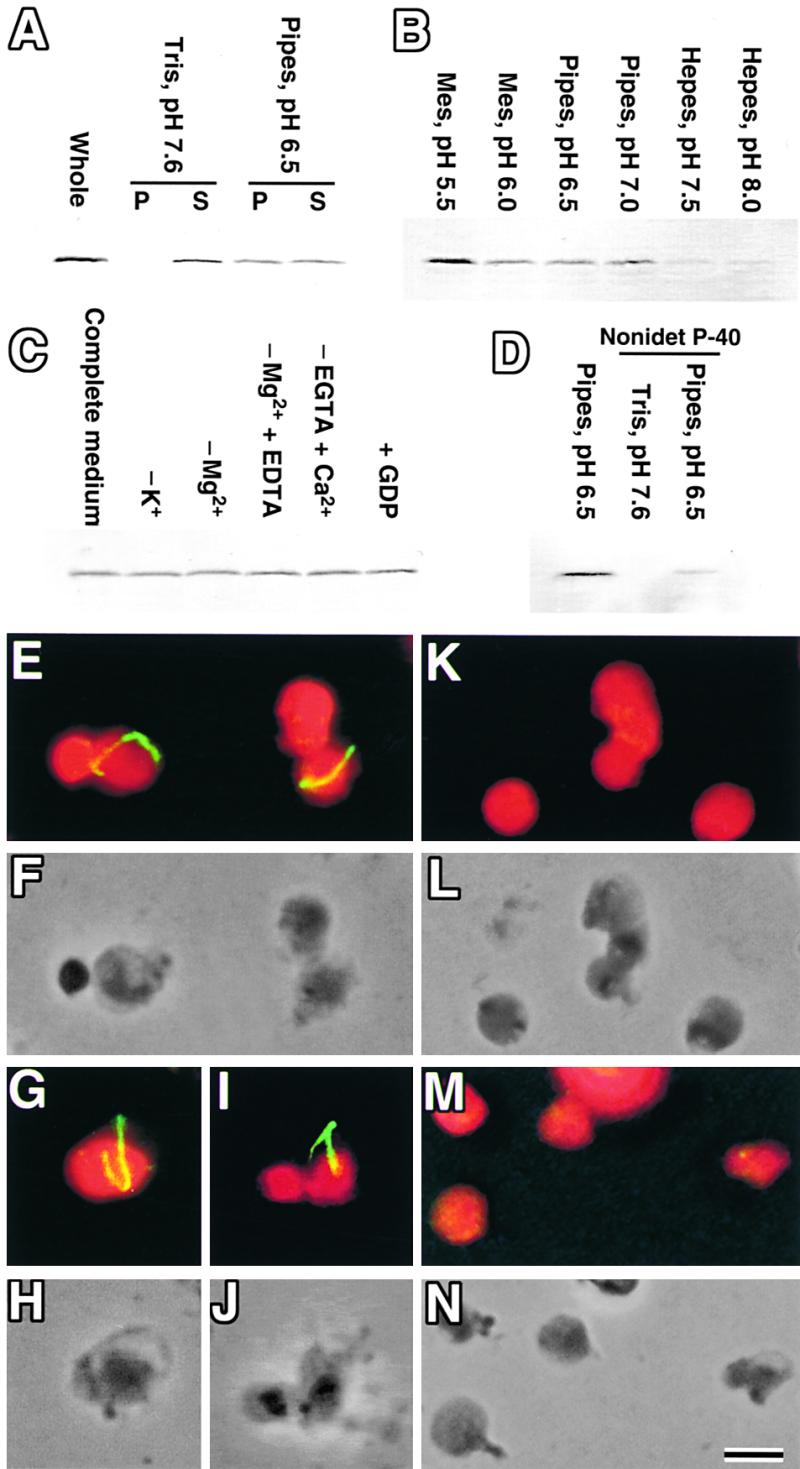
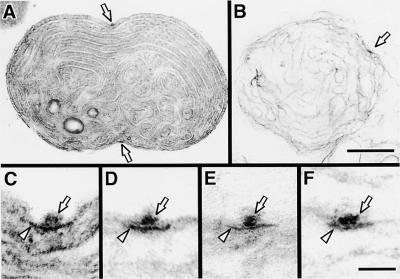
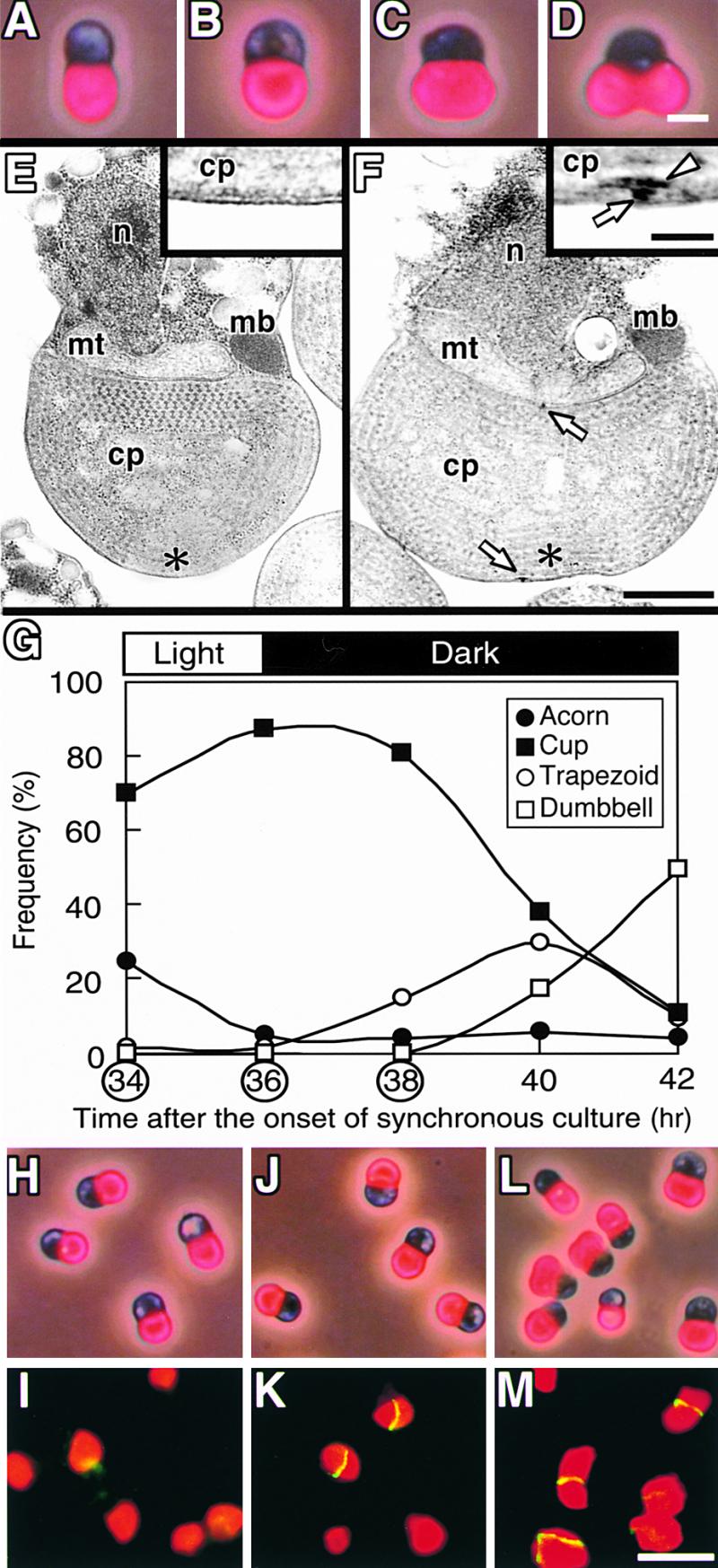
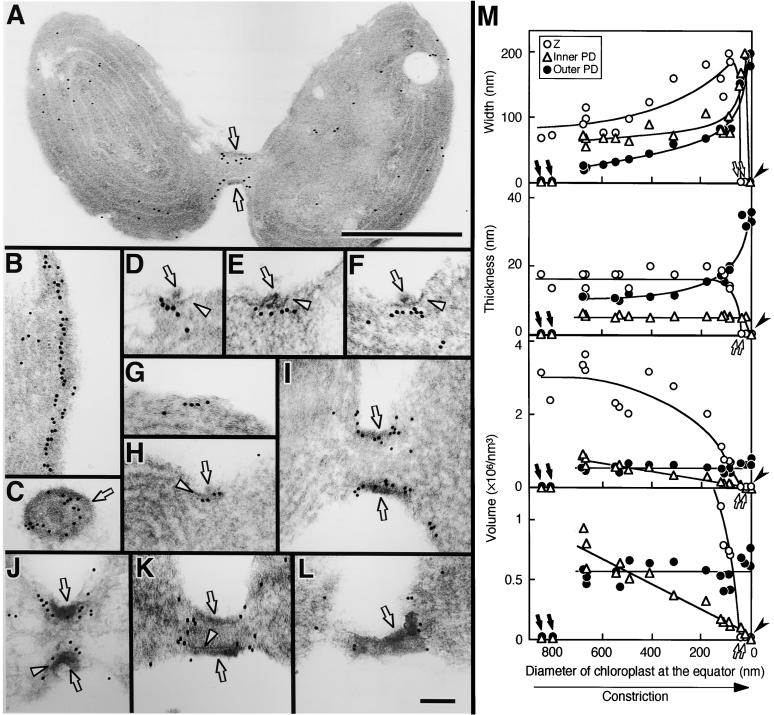
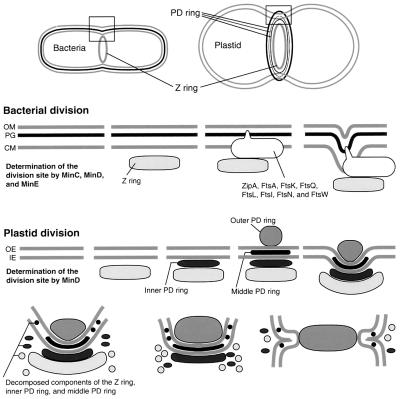
During plastid division, two structures have been detected at the division site in separate analyses. The plastid-dividing ring can be detected by transmission electron microscopy as two (or three) electron-dense rings: an outer ring on the cytosolic face of the outer envelope, occasionally a middle ring in the intermembrane space, and an inner ring on the stromal face of the inner envelope. The FtsZ ring, which plays a central role in bacterial division, also is involved in plastid division and is believed to have descended to plastids from cyanobacterial endosymbiosis. The relationship between the two structures is not known, although there is discussion regarding whether they are identical. Biochemical and immunocytochemical investigations, using synchronized chloroplasts of the red alga Cyanidioschyzon merolae, showed that the plastid FtsZ ring is distinct and separable from the plastid-dividing ring. The FtsZ ring localizes in stroma and faces the inner plastid-dividing ring at the far side from the inner envelope. The FtsZ ring and the inner and outer plastid-dividing rings form in that order before plastid division. The FtsZ ring disappears at the late stage of constriction before dissociation of the plastid-dividing ring, when the constriction is still in progress. Our results suggest that the FtsZ ring;-based system, which originated from a plastid ancestor, cyanobacteria, and the plastid-dividing ring;-based system, which probably originated from host eukaryotic cells, form a complex and are involved in plastid division by distinct modes.
Figures
References
-
- Beech, P.L., and Gilson, P.R. (2000). FtsZ and organelle division in protists. Protist 151, 11–16. - PubMed
-
- Beech, P.L., Nheu, T., Schultz, T., Herbert, S., Lithgow, T., Gilson, P.R., and McFadden, G.I. (2000). Mitochondrial FtsZ in a chromophyte alga. Science 287, 1276–1279. - PubMed
-
- Bramhill, D. (1997). Bacterial cell division. Annu. Rev. Cell Dev. Biol. 13, 395–424. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
