

Cdc42p functions at the docking stage of yeast vacuole membrane fusion
- PMID: 11598009
- PMCID: PMC125663
- DOI: 10.1093/emboj/20.20.5657
Cdc42p functions at the docking stage of yeast vacuole membrane fusion
Abstract
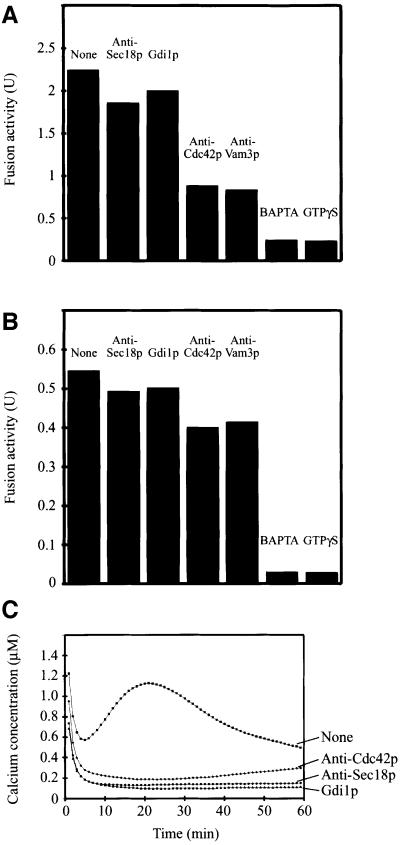
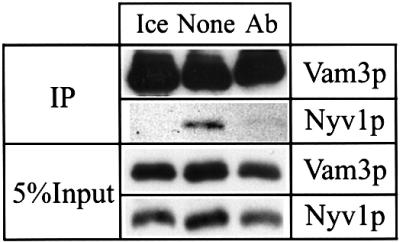
Membrane fusion reactions have been considered to be primarily regulated by Rab GTPases. In the model system of homotypic vacuole fusion in the yeast Saccharomyces cerevisiae, we show that Cdc42p, a member of the Rho family of GTPases, has a direct role in membrane fusion. Genetic evidence suggested a relationship between Cdc42p and Vtc1p/Nrf1p, a central part of the vacuolar membrane fusion machinery. Vacuoles from cdc42 temperature-sensitive mutants are deficient for fusion at the restrictive temperature. Specific amino acid changes on the Cdc42p protein surface in these mutants define the putative interaction domain that is crucial for its function in membrane fusion. Affinity-purified antibodies to this domain inhibited the in vitro fusion reaction. Using these antibodies in kinetic analyses and assays for subreactions of the priming, docking and post-docking phase of the reaction, we show that Cdc42p action follows Ypt7p-dependent tethering, but precedes the formation of trans-SNARE complexes. Thus, our data define an effector binding domain of Cdc42p by which it regulates the docking reaction of vacuole fusion.
Figures
Similar articles
-
Rho1p and Cdc42p act after Ypt7p to regulate vacuole docking.EMBO J. 2001 Oct 15;20(20):5650-6. doi: 10.1093/emboj/20.20.5650. EMBO J. 2001. PMID: 11598008 Free PMC article.
-
A soluble SNARE drives rapid docking, bypassing ATP and Sec17/18p for vacuole fusion.EMBO J. 2004 Jul 21;23(14):2765-76. doi: 10.1038/sj.emboj.7600286. Epub 2004 Jul 8. EMBO J. 2004. PMID: 15241469 Free PMC article.
-
The N-terminal domain of the t-SNARE Vam3p coordinates priming and docking in yeast vacuole fusion.Mol Biol Cell. 2001 Nov;12(11):3375-85. doi: 10.1091/mbc.12.11.3375. Mol Biol Cell. 2001. PMID: 11694574 Free PMC article.
-
Membrane fusion: five lipids, four SNAREs, three chaperones, two nucleotides, and a Rab, all dancing in a ring on yeast vacuoles.Annu Rev Cell Dev Biol. 2010;26:115-36. doi: 10.1146/annurev-cellbio-100109-104131. Annu Rev Cell Dev Biol. 2010. PMID: 20521906 Review.
-
Saccharomyces cerevisiae--a model organism for the studies on vacuolar transport.Acta Biochim Pol. 2001;48(4):1025-42. Acta Biochim Pol. 2001. PMID: 11995965 Review.
Cited by
-
Hierarchy of protein assembly at the vertex ring domain for yeast vacuole docking and fusion.J Cell Biol. 2003 Feb 3;160(3):365-74. doi: 10.1083/jcb.200209095. J Cell Biol. 2003. PMID: 12566429 Free PMC article.
-
Exo-endocytic trafficking and the septin-based diffusion barrier are required for the maintenance of Cdc42p polarization during budding yeast asymmetric growth.Mol Biol Cell. 2011 Mar 1;22(5):624-33. doi: 10.1091/mbc.E10-06-0484. Epub 2011 Jan 5. Mol Biol Cell. 2011. PMID: 21209323 Free PMC article.
-
FGD2, a CDC42-specific exchange factor expressed by antigen-presenting cells, localizes to early endosomes and active membrane ruffles.J Biol Chem. 2008 Dec 5;283(49):34002-12. doi: 10.1074/jbc.M803957200. Epub 2008 Oct 6. J Biol Chem. 2008. PMID: 18838382 Free PMC article.
-
Vtc5, a Novel Subunit of the Vacuolar Transporter Chaperone Complex, Regulates Polyphosphate Synthesis and Phosphate Homeostasis in Yeast.J Biol Chem. 2016 Oct 14;291(42):22262-22275. doi: 10.1074/jbc.M116.746784. Epub 2016 Sep 1. J Biol Chem. 2016. PMID: 27587415 Free PMC article.
-
The Vtc proteins in vacuole fusion: coupling NSF activity to V(0) trans-complex formation.EMBO J. 2002 Feb 1;21(3):259-69. doi: 10.1093/emboj/21.3.259. EMBO J. 2002. PMID: 11823419 Free PMC article.
References
-
- Abdul-Manan N., Aghazadeh,B., Liu,G.A., Majumdar,A., Ouerfelli,O., Siminovitch,K.A. and Rosen,M.K. (1999) Structure of Cdc42 in complex with the GTPase-binding domain of the ‘Wiskott–Aldrich syndrome’ protein. Nature, 399, 379–383. - PubMed
-
- Ayscough K.R. (2000) Endocytosis and the development of cell polarity in yeast require a dynamic F-actin cytoskeleton. Curr. Biol., 10, 1587–1590. - PubMed
-
- Bernstein B.W., DeWitt,M. and Bamburg,J.R. (1998) Actin disassembles reversibly during electrically induced recycling of synaptic vesicles in cultured neurons. Brain Res. Mol. Brain Res., 53, 236–251. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
