

Administration of superantigens protects mice from lethal Listeria monocytogenes infection by enhancing cytotoxic T cells
- PMID: 11598032
- PMCID: PMC100037
- DOI: 10.1128/IAI.69.11.6633-6642.2001
Administration of superantigens protects mice from lethal Listeria monocytogenes infection by enhancing cytotoxic T cells
Abstract
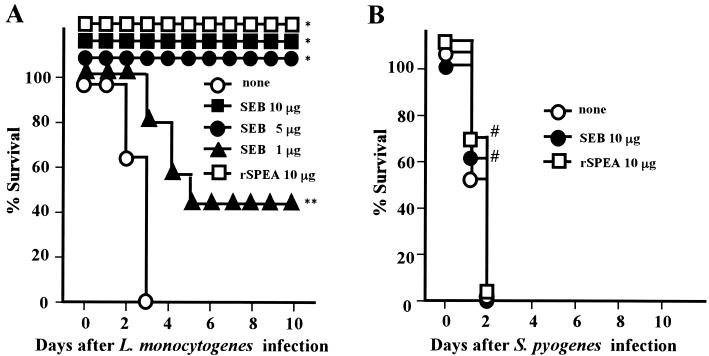
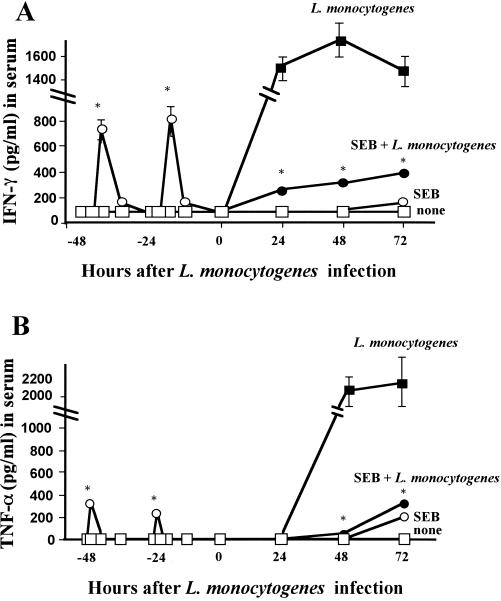
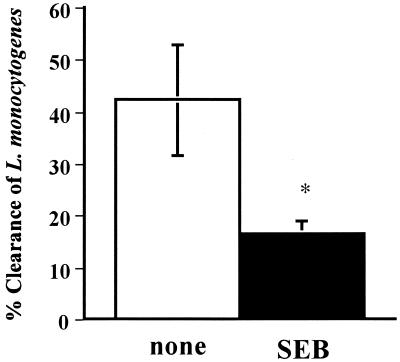
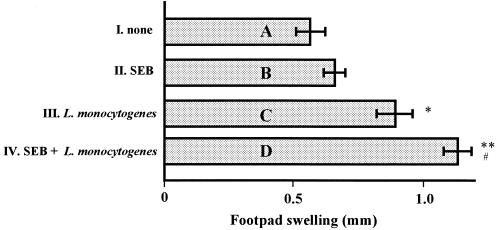
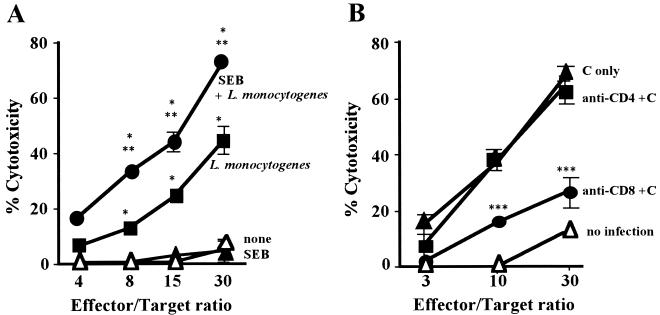
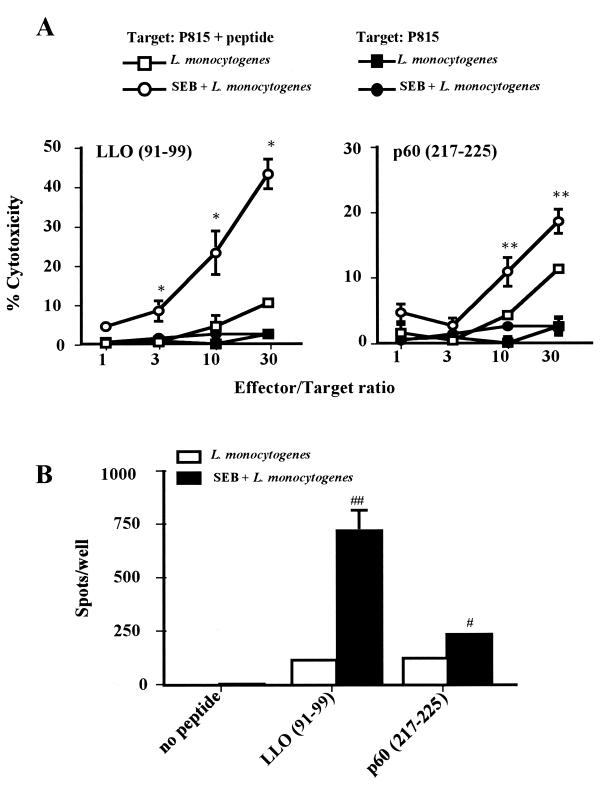
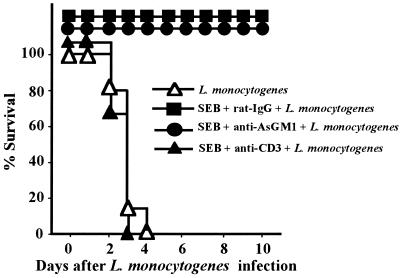
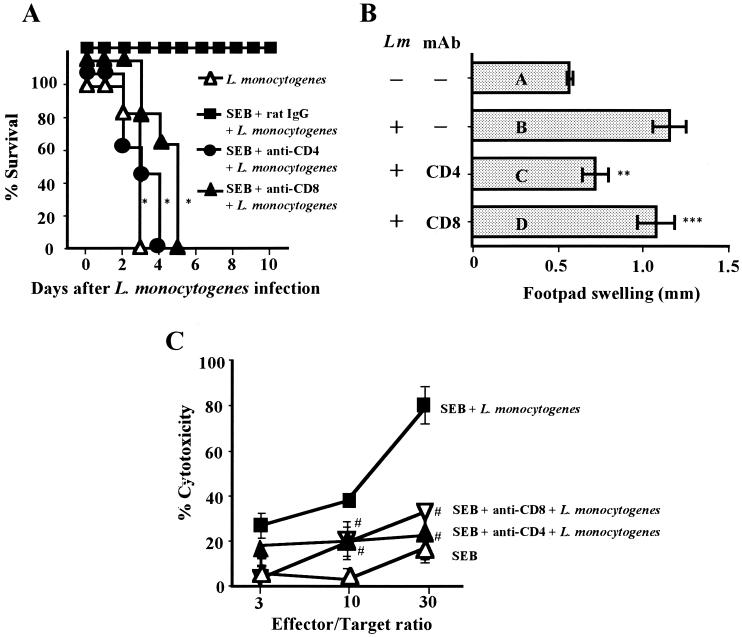
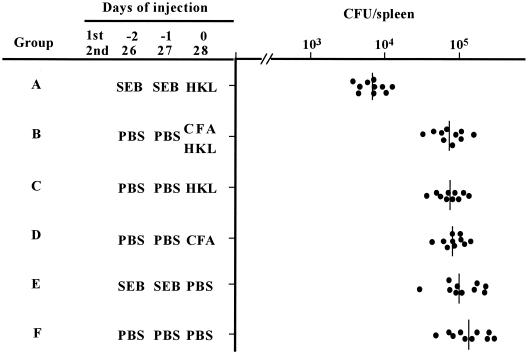
Superantigens stimulate T-cell-receptor Vbeta-selective T-cell proliferation accompanying the release of cytokines, which may eventually protect the host from microbial infections. We investigated here whether superantigens can rescue the host from lethal bacterial infection. Mice were pretreated with Staphylococcus aureus enterotoxin B (SEB) 1 and 2 days before bacterial infection, and the mortality of infected mice was assessed. SEB pretreatment protected mice from lethal infection with Listeria monocytogenes but not from lethal infection with Streptococcus pyogenes. This enhanced protection was also observed upon pretreatment with recombinant streptococcal pyrogenic exotoxin A. Furthermore, L. monocytogenes-specific delayed-type hypersensitivity (DTH) due to type 1 helper T (Th1) cells and the cytotoxicity of CD8(+) T cells were significantly enhanced after SEB administration and bacterial infection. Depletion of either CD4(+) T cells or CD8(+) T cells in SEB-pretreated mice completely abolished this protection. This phenomenon was ascribed to the elimination of L. monocytogenes-specific CD8(+) cytotoxic T lymphocytes (CTL). It was found that CD4(+) T cells contributed to the induction of the CTL populations. Furthermore, SEB pretreatment of heat-killed L. monocytogenes-immunized mice enhanced the protection from challenge of L. monocytogenes. Taken together, these results indicated that administrations of superantigens protected mice from infection with L. monocytogenes, which was dependent on the enhanced L. monocytogenes-specific CTL activity in the presence of CD4(+) T cells, and superantigens exhibited adjuvant activity in the immunization against intracellular pathogens.
Figures
References
-
- Bancroft G J, Sheehan K C F, Schreiber R D, Unanue E R. Tumor necrosis factor is involved in the T cell-independent pathway of macrophage activation in scid mice. J Immunol. 1989;143:127–130. - PubMed
-
- Blackman M A, Woodland D L. In vivo effects of superantigens. Life Sci. 1995;57:1717–1735. - PubMed
-
- Buller R M L, Holmes K L, Hügin A, Frederickson T N, Morse H C., III Induction of cytotoxic T-cell responses in vivo in the absence of CD4 helper cells. Nature. 1987;328:77–79. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
