

Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need?
- PMID: 11599559
- PMCID: PMC434399
- DOI: 10.1379/1466-1268(2001)006<0177:aathst>2.0.co;2
Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need?
Abstract
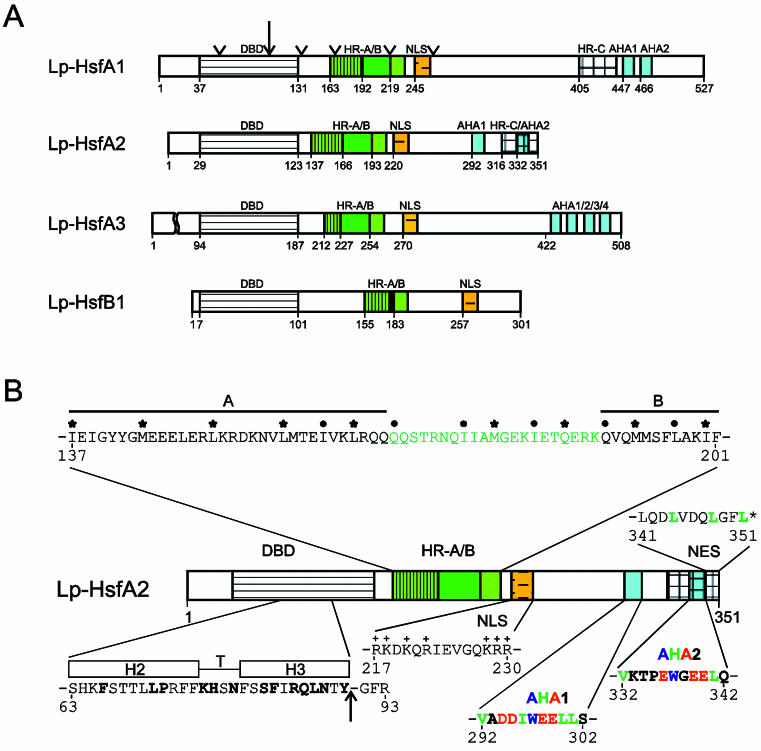
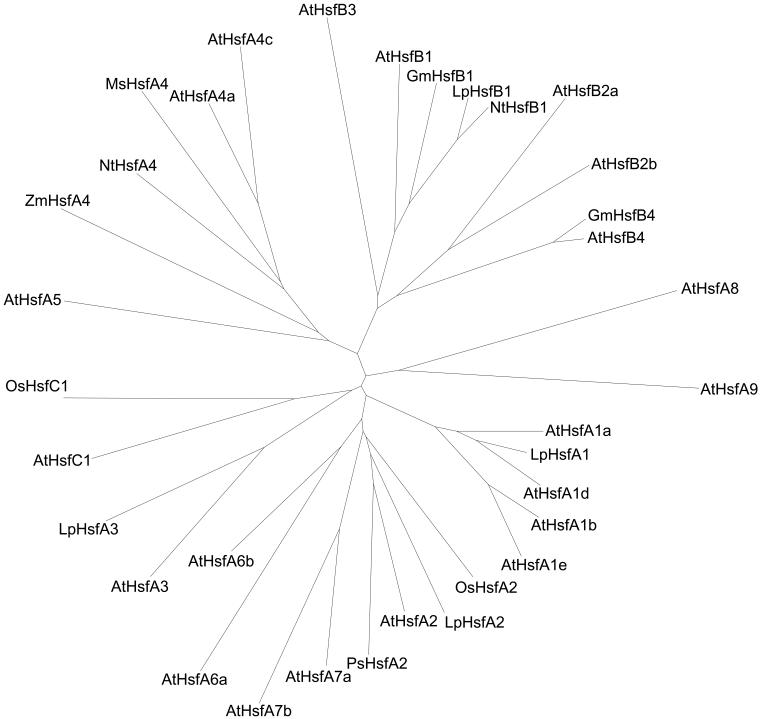
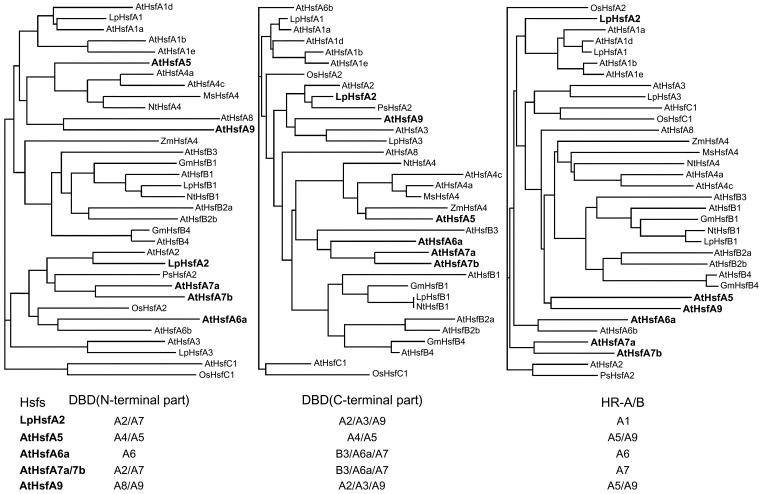
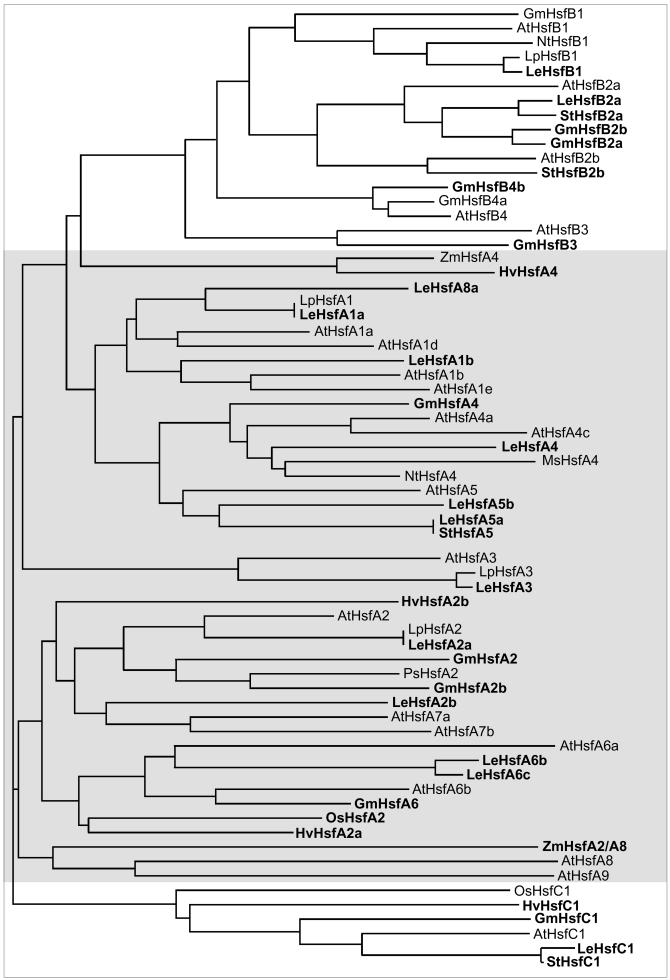
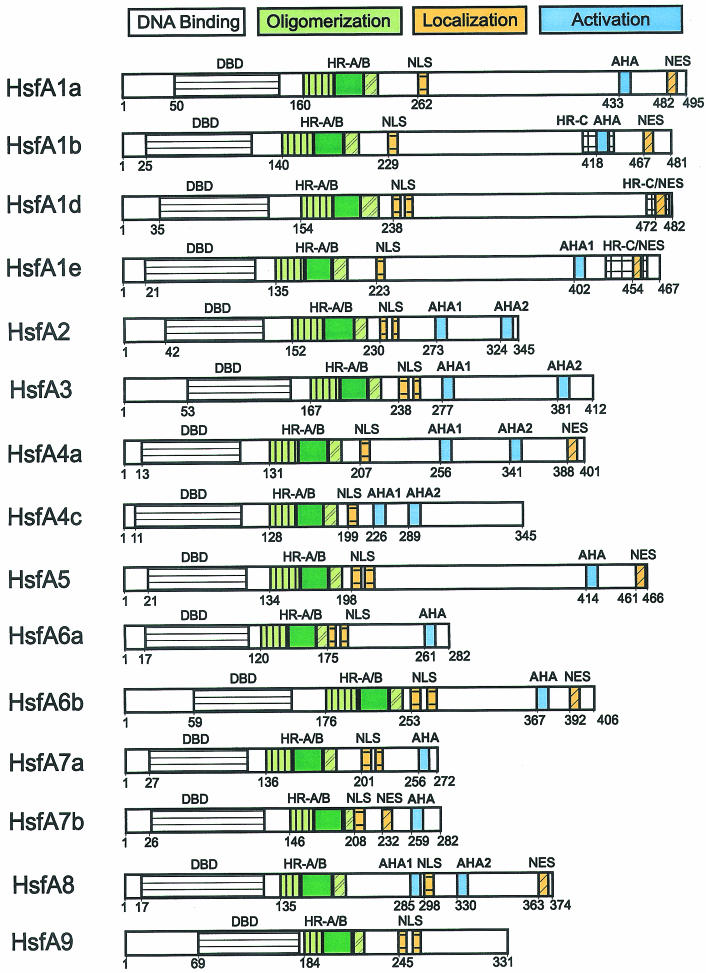
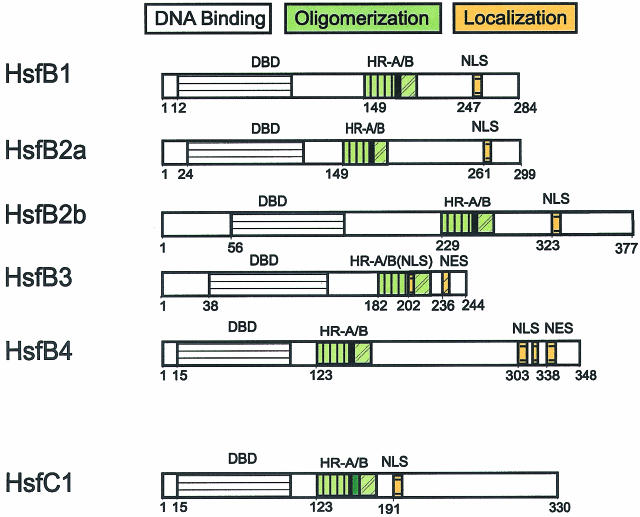
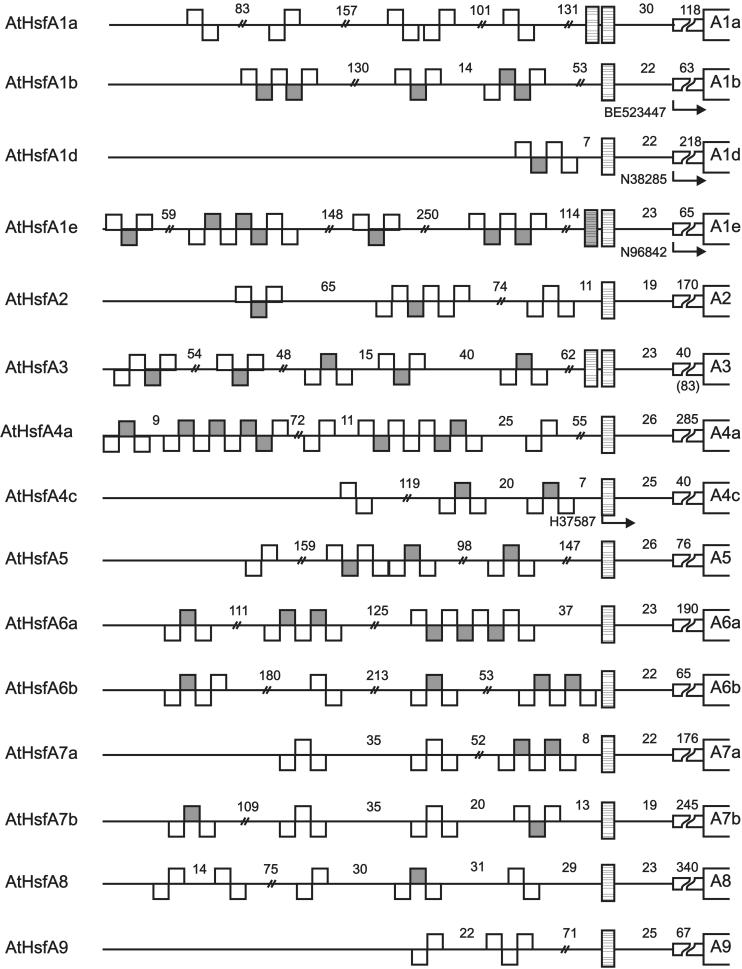
Sequencing of the Arabidopsis genome revealed a unique complexity of the plant heat stress transcription factor (Hsf) family. By structural characteristics and phylogenetic comparison, the 21 representatives are assigned to 3 classes and 14 groups. Particularly striking is the finding of a new class of Hsfs (AtHsfC1) closely related to Hsf1 from rice and to Hsfs identified from frequently found expressed sequence tags of tomato, potato, barley, and soybean. Evidently, this new type of Hsf is well expressed in different plant tissues. Besides the DNA binding and oligomerization domains (HR-A/B region), we identified other functional modules of Arabidopsis Hsfs by sequence comparison with the well-characterized tomato Hsfs. These are putative motifs for nuclear import and export and transcriptional activation (AHA motifs). There is intriguing flexibility of size and sequence in certain parts of the otherwise strongly conserved N-terminal half of these Hsfs. We have speculated about possible exon-intron borders in this region in the ancient precursor gene of plant Hsfs, similar to the exon-intron structure of the present mammalian Hsf-encoding genes.
Figures

References
-
- Aranda MA, Escaler M, Thomas CL, Maule AJ. A heat stress transcription factor in pea is differentially controlled by heat and virus replication. Plant J. 1999;20:153–161. - PubMed
-
- Bharti K, Schmidt E, Lyck R, Bublak D, Scharf KD. Isolation and characterization of HsfA3, a new heat stress transcription factor of Lycopersicon peruvianum. Plant J. 2000;22:355–365. - PubMed
-
- Bienz M, Pelham HRB. Mechanisms of heat-shock gene activation in higher eukaryotes. Adv Genet. 1987;24:31–72. - PubMed
-
- Clos J, Westwood JT, Becker PB, Wilson S, Lambert U, Wu C. Molecular cloning and expression of a heaxameric Drosophila heat stress factor subject to negative regulation. Cell. 1990;63:1085–1097. - PubMed
-
- Czarnecka-Verner E, Yuan CX, Fox PC, Gurley WB. Isolation and characterization of six heat stress transcription factor cDNA clones from soybean. Plant Mol Biol. 1995;29:37–51. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases