

The role of acetylation in rDNA transcription
- PMID: 11600700
- PMCID: PMC60214
- DOI: 10.1093/nar/29.20.4114
The role of acetylation in rDNA transcription
Abstract
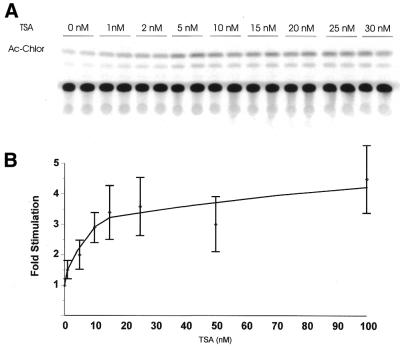
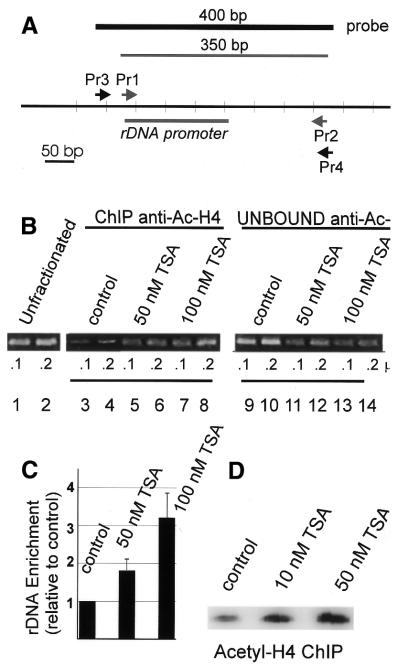
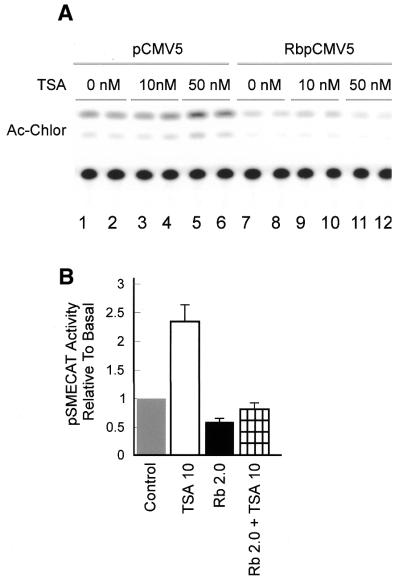
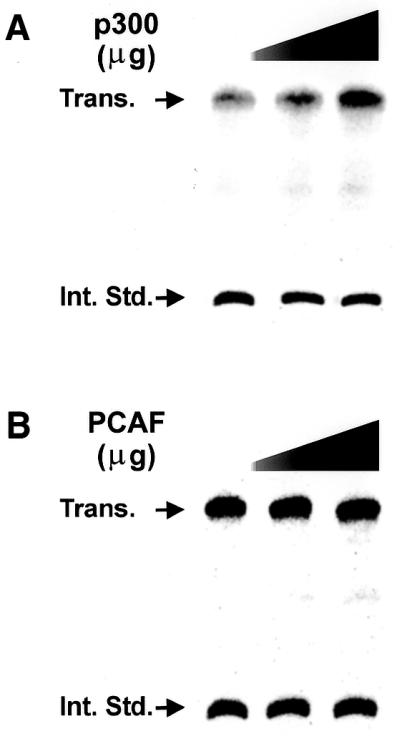
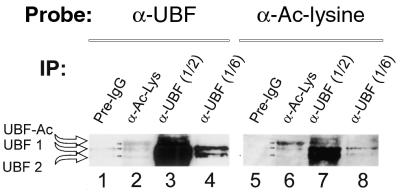
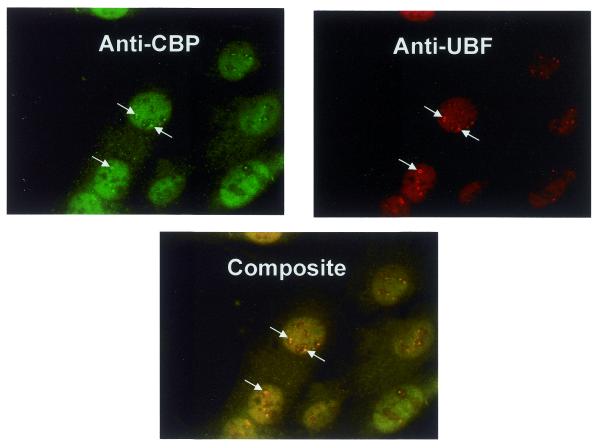
Treatment of NIH 3T3 cells with trichostatin A (TSA), an inhibitor of histone deacetylase (HDAC), resulted in a dose-dependent increase in transcription from a rDNA reporter and from endogenous rRNA genes. Chromatin immunoprecipitation using anti-acetyl-histone H4 antibodies demonstrated a direct effect of TSA on the acetylation state of the ribosomal chromatin. TSA did not reverse inhibition of transcription from the rDNA reporter by retinoblastoma (Rb) protein, suggesting that the main mechanism by which Rb blocks rDNA transcription may not involve recruitment of deacetylases to rDNA chromatin. Overexpression of histone transacetylases p300, CBP and PCAF stimulated transcription in transfected NIH 3T3 cells. Recombinant p300, but not PCAF, stimulated rDNA transcription in vitro in the absence of nucleosomes, suggesting that the stimulation of rDNA transcription by TSA might have a chromatin-independent component. We found that the rDNA transcription factor UBF was acetylated in vivo. Finally, we also demonstrated the nucleolar localization of CBP. Our results suggest that the organization of ribosomal chromatin of higher eukaryotes is not static and that acetylation may be involved in affecting these dynamic changes directly through histone acetylation and/or through acetylation of UBF or one of the other components of rDNA transcription.
Figures


Similar articles
-
Competitive recruitment of CBP and Rb-HDAC regulates UBF acetylation and ribosomal transcription.Mol Cell. 2000 Nov;6(5):1059-66. doi: 10.1016/s1097-2765(00)00104-0. Mol Cell. 2000. PMID: 11106745
-
Inhibition of MMTV transcription by HDAC inhibitors occurs independent of changes in chromatin remodeling and increased histone acetylation.Oncogene. 2003 Jul 31;22(31):4807-18. doi: 10.1038/sj.onc.1206722. Oncogene. 2003. PMID: 12894222
-
Trichostatin A induces transforming growth factor beta type II receptor promoter activity and acetylation of Sp1 by recruitment of PCAF/p300 to a Sp1.NF-Y complex.J Biol Chem. 2005 Mar 18;280(11):10047-54. doi: 10.1074/jbc.M408680200. Epub 2005 Jan 12. J Biol Chem. 2005. PMID: 15647279
-
Acetylation of TAF(I)68, a subunit of TIF-IB/SL1, activates RNA polymerase I transcription.EMBO J. 2001 Mar 15;20(6):1353-62. doi: 10.1093/emboj/20.6.1353. EMBO J. 2001. PMID: 11250901 Free PMC article.
-
Chromatin: linking structure and function in the nucleolus.Chromosoma. 2009 Feb;118(1):11-23. doi: 10.1007/s00412-008-0184-2. Epub 2008 Oct 17. Chromosoma. 2009. PMID: 18925405 Review.
Cited by
-
ESET methylates UBF at K232/254 and regulates nucleolar heterochromatin plasticity and rDNA transcription.Nucleic Acids Res. 2014 Feb;42(3):1628-43. doi: 10.1093/nar/gkt1041. Epub 2013 Nov 14. Nucleic Acids Res. 2014. PMID: 24234436 Free PMC article.
-
ING1 regulates rRNA levels by altering nucleolar chromatin structure and mTOR localization.Nucleic Acids Res. 2017 Feb 28;45(4):1776-1792. doi: 10.1093/nar/gkw1161. Nucleic Acids Res. 2017. PMID: 27903908 Free PMC article.
-
FGFR2 mutations in bent bone dysplasia syndrome activate nucleolar stress and perturb cell fate determination.Hum Mol Genet. 2017 Sep 1;26(17):3253-3270. doi: 10.1093/hmg/ddx209. Hum Mol Genet. 2017. PMID: 28595297 Free PMC article.
-
Characterization of the fission yeast ribosomal DNA binding factor: components share homology with Upstream Activating Factor and with SWI/SNF subunits.Nucleic Acids Res. 2002 Dec 15;30(24):5347-59. doi: 10.1093/nar/gkf683. Nucleic Acids Res. 2002. PMID: 12490702 Free PMC article.
-
The coordinated management of ribosome and translation during injury and regeneration.Front Cell Dev Biol. 2023 Jun 22;11:1186638. doi: 10.3389/fcell.2023.1186638. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37427381 Free PMC article. Review.
References
-
- Moss T. and Stefanovsky,V.Y. (1995) Promotion and regulation of ribosomal transcription in eukaryota by RNA polymerase I. Prog. Nucleic Acid Res. Mol. Biol., 50, 25–66. - PubMed
-
- Hannan K., Hannan,R.D. and Rothblum,L.I. (1998) Transcription by RNA polymerase I. Front. Biosci., 3, d376–d398. - PubMed
-
- Paule M.R. (1998) Transcription of Ribosomal RNA Genes by Eucaryotic RNA Polymerase I. Springer-Verlag and R.G. Landes, Georgetown, TX.
-
- Reeder R.H. (1999) Regulation of RNA polymerase I transcription in yeast and vertebrates. Prog. Nucleic Acid Res. Mol. Biol., 62, 293–327. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
