

A DAP12-mediated pathway regulates expression of CC chemokine receptor 7 and maturation of human dendritic cells
- PMID: 11602640
- PMCID: PMC2193511
- DOI: 10.1084/jem.194.8.1111
A DAP12-mediated pathway regulates expression of CC chemokine receptor 7 and maturation of human dendritic cells
Abstract
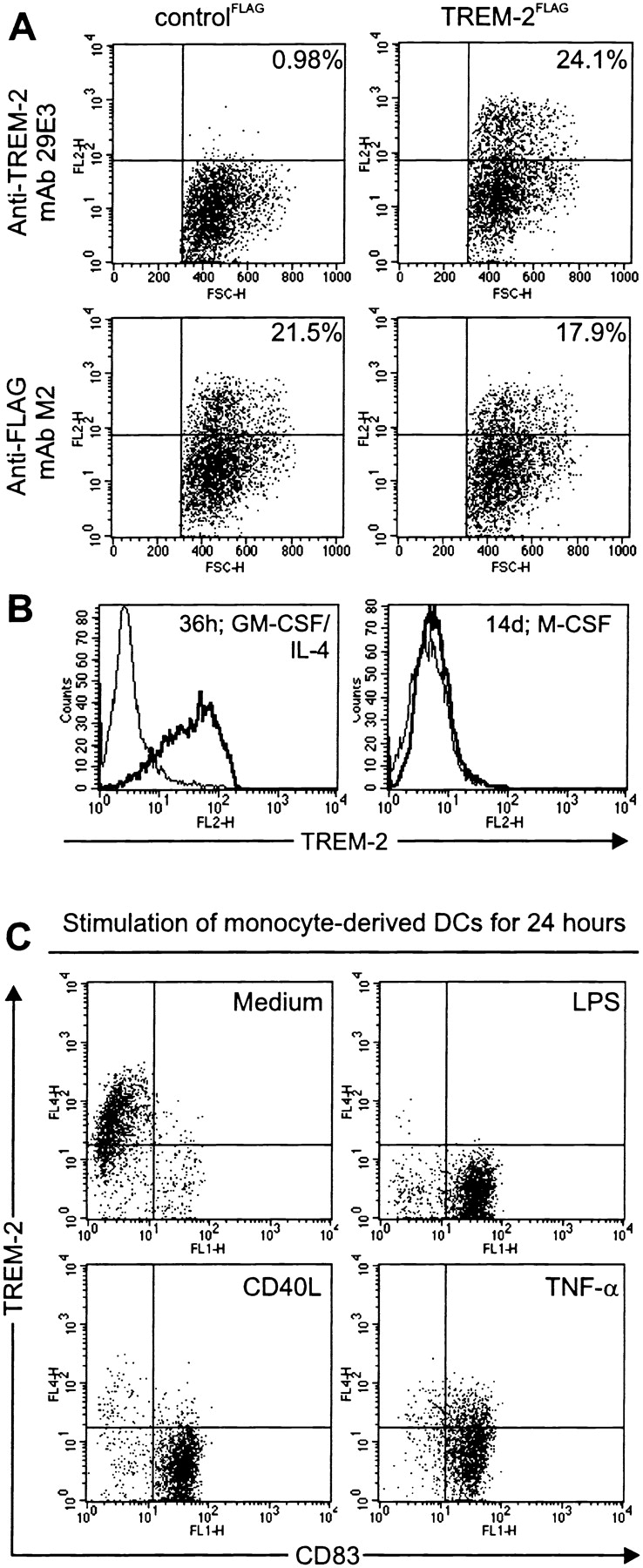
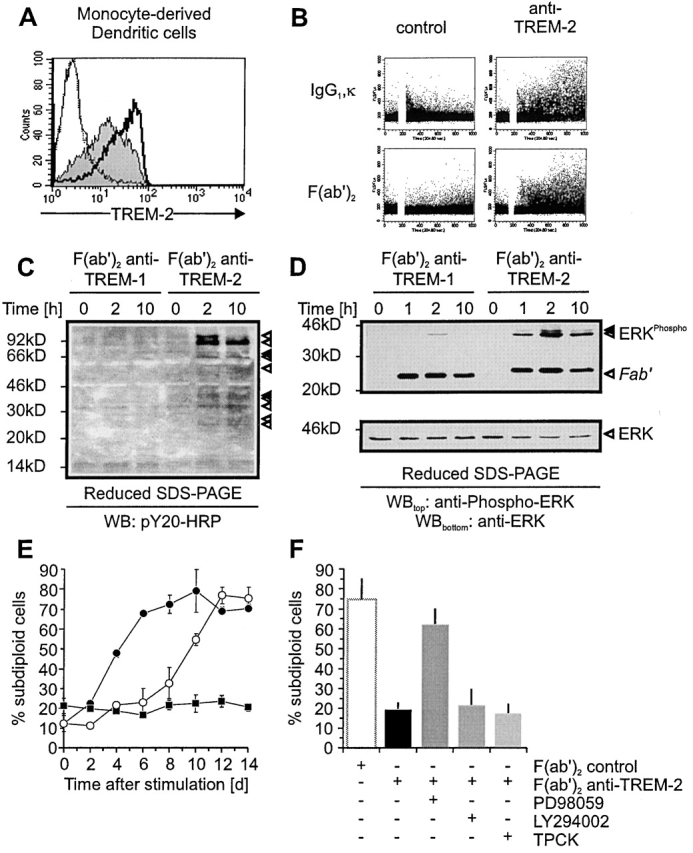
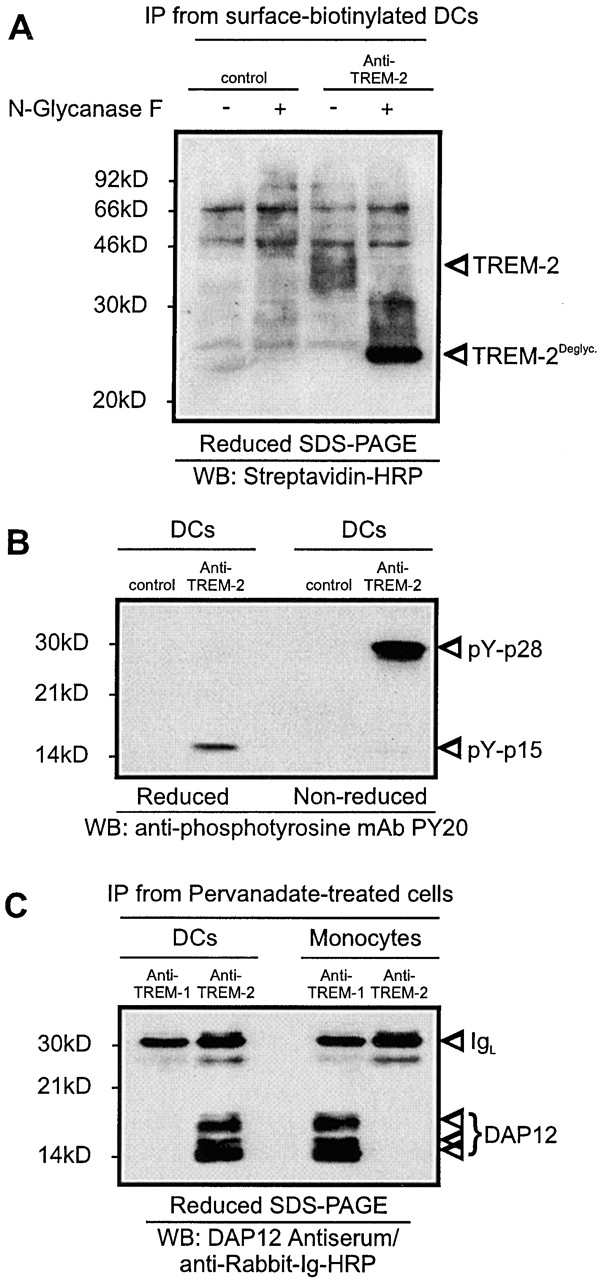
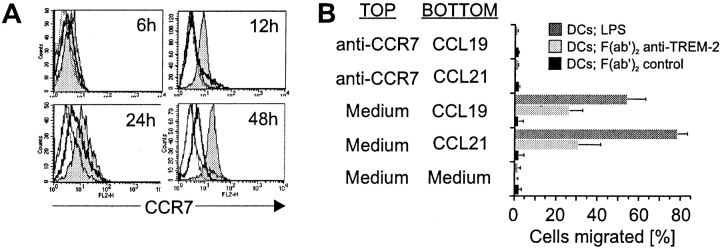
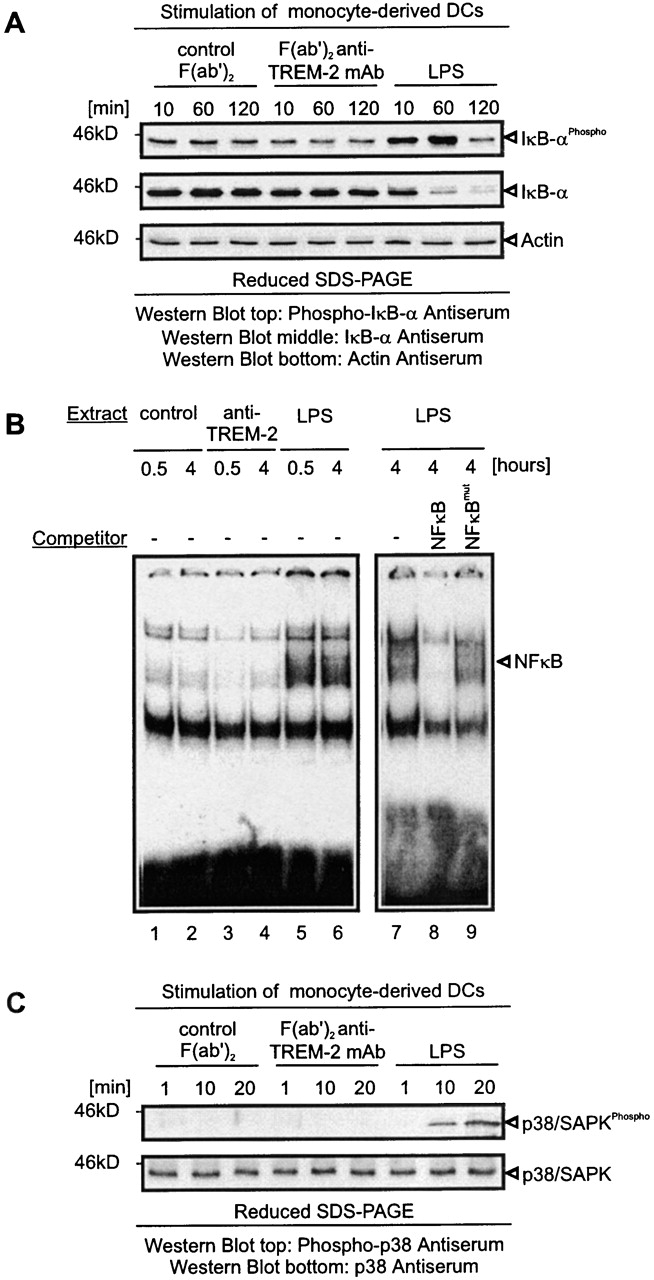
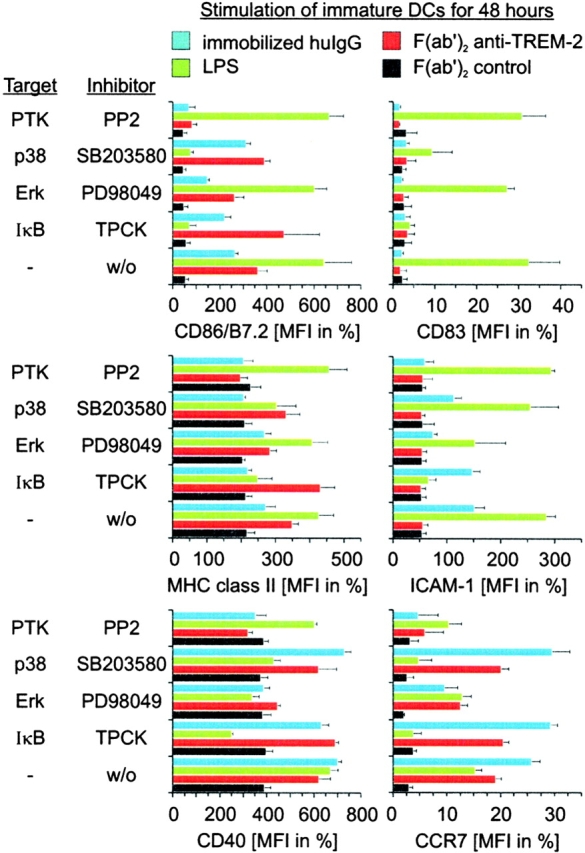
Gene targeting of the adaptor molecule DAP12 in mice caused abnormal distribution and impaired antigen presentation capacity of dendritic cells (DCs). However, the DAP12-associated receptors expressed on DCs and their functions have not been identified yet. Here we show that the triggering receptor expressed on myeloid cells-2 (TREM-2) is a cell surface receptor on human monocyte-derived DCs, which is associated with DAP12. TREM-2/DAP12 promotes upregulation of CC chemokine receptor 7, partial DC maturation, and DC survival through activation of protein tyrosine kinases and extracellular signal-regulated kinase. In contrast to Toll-like receptor-mediated signaling, TREM2/DAP12 stimulation is independent of nuclear factor-kappaB and p38 stress-activated protein kinase. This novel DC activation pathway may regulate DC homeostasis and amplify DC responses to pathogens, explaining the phenotype observed in DAP12-deficient mice.
Figures
References
-
- Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
-
- Gallucci S., Lolkema M., Matzinger P. Natural adjuvantsendogenous activators of dendritic cells. Nat. Med. 1999;11:1249–1255. - PubMed
-
- Sozzani S., Allavena P., D'Amico G., Luini W., Bianchi G., Kataura M., Imai T., Yoshie O., Bonecchi R., Mantovani A. Differential regulation of chemokine receptors during dendritic cell maturationa model for their trafficking properties. J. Immunol. 1998;161:1083–1086. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
