

An attenuating mutation in the 2A protease of swine vesicular disease virus, a picornavirus, regulates cap- and internal ribosome entry site-dependent protein synthesis
- PMID: 11602706
- PMCID: PMC114646
- DOI: 10.1128/JVI.75.22.10643-10650.2001
An attenuating mutation in the 2A protease of swine vesicular disease virus, a picornavirus, regulates cap- and internal ribosome entry site-dependent protein synthesis
Abstract
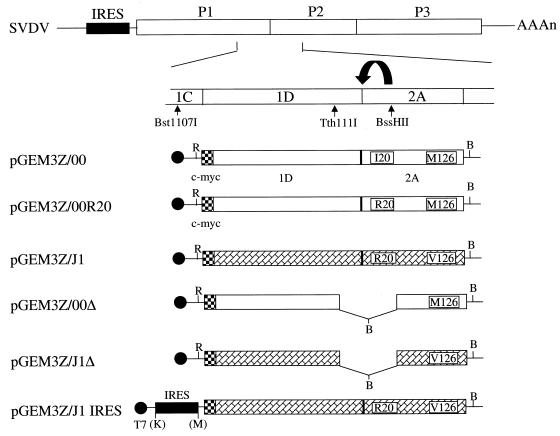
Virulent and avirulent strains of swine vesicular disease virus (SVDV), a picornavirus, have been characterized previously. The major determinants for attenuation have been mapped to specific residues in the 1D-2A-coding region. The properties of the 2A proteases from the virulent and avirulent strains of SVDV have now been examined. Both proteases efficiently cleaved the 1D/2A junction in vitro and in vivo. However, the 2A protease of the avirulent strain of SVDV was much less effective than the virulent-virus 2A protease at inducing cleavage of translation initiation factor eIF4GI within transfected cells. Hence the virulent-virus 2A protease is much more effective at inhibiting cap-dependent protein synthesis. Furthermore, the virulent-virus 2A protease strongly stimulated the internal ribosome entry sites (IRESs) from coxsackievirus B4 and from SVDV, while the avirulent-virus 2A protease was significantly less active in these assays. Thus, the different properties of the 2A proteases from the virulent and avirulent strains of SVDV in regulating protein synthesis initiation reflect the distinct pathogenic properties of the viruses from which they are derived. A single amino acid substitution, adjacent to His21 of the catalytic triad, is sufficient to confer the characteristics of the virulent-strain 2A protease on the avirulent-strain protease. It is concluded that the efficiency of picornavirus protein synthesis, controlled directly by the IRES or indirectly by the 2A protease, can determine virus virulence.
Figures
References
-
- Belsham G J, Jackson R J. Translation initiation on picornavirus RNA. In: Sonenberg N, Hershey J W B, Mathews M B, editors. Translational control of gene expression. Monograph 39. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 2000. pp. 869–900.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
