

C terminus of infectious bursal disease virus major capsid protein VP2 is involved in definition of the T number for capsid assembly
- PMID: 11602723
- PMCID: PMC114663
- DOI: 10.1128/JVI.75.22.10815-10828.2001
C terminus of infectious bursal disease virus major capsid protein VP2 is involved in definition of the T number for capsid assembly
Abstract
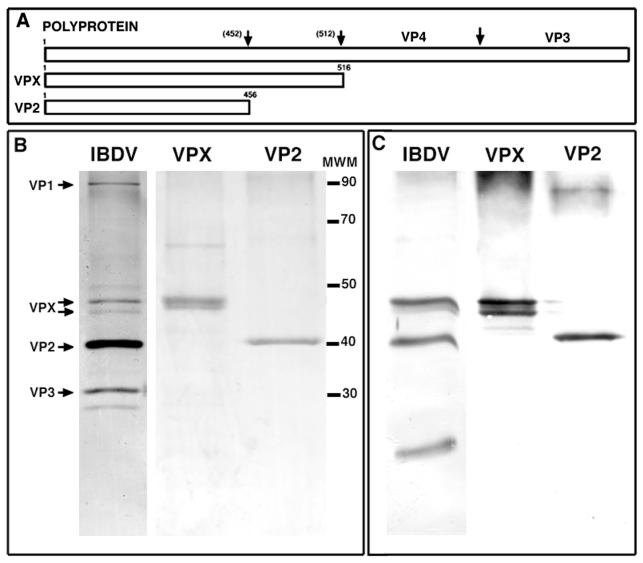
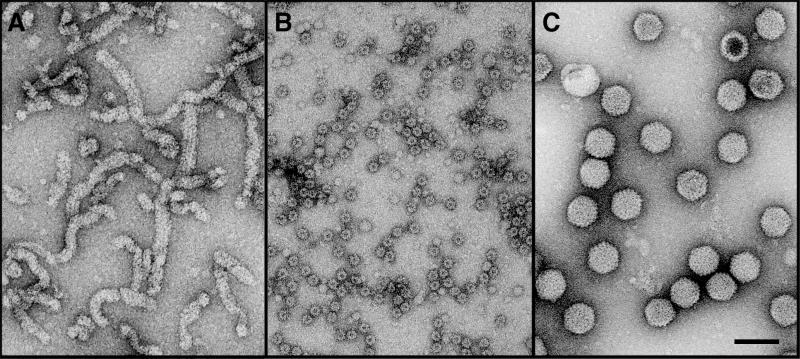
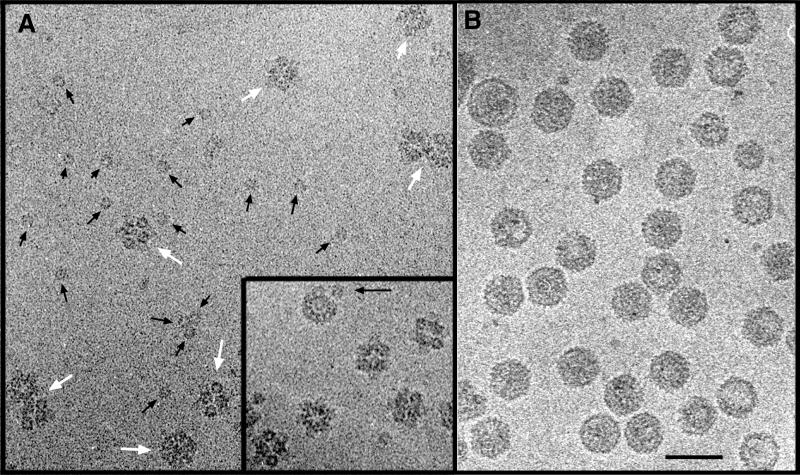
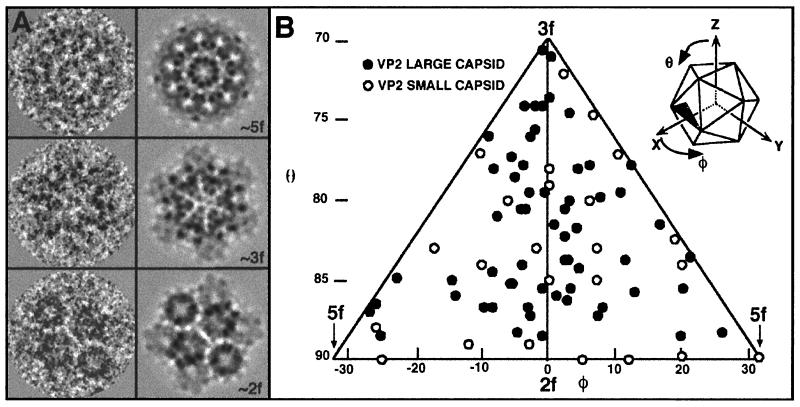
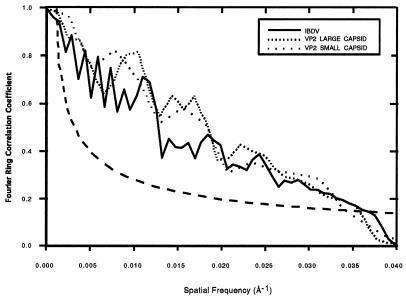
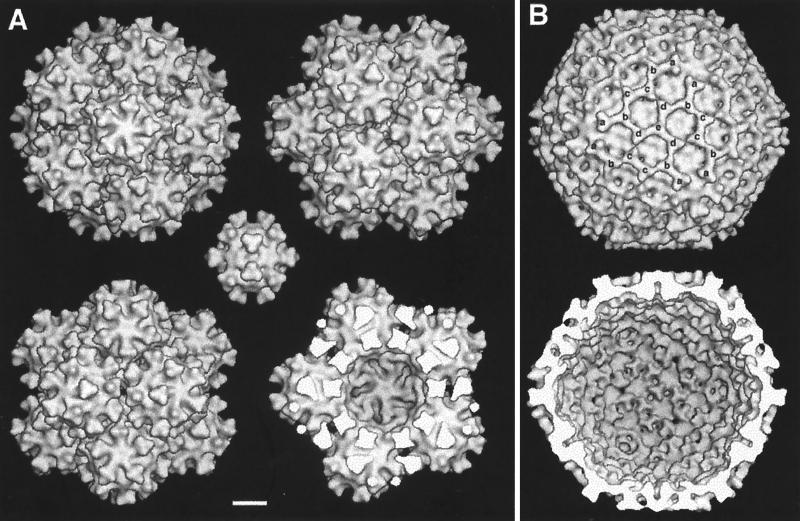
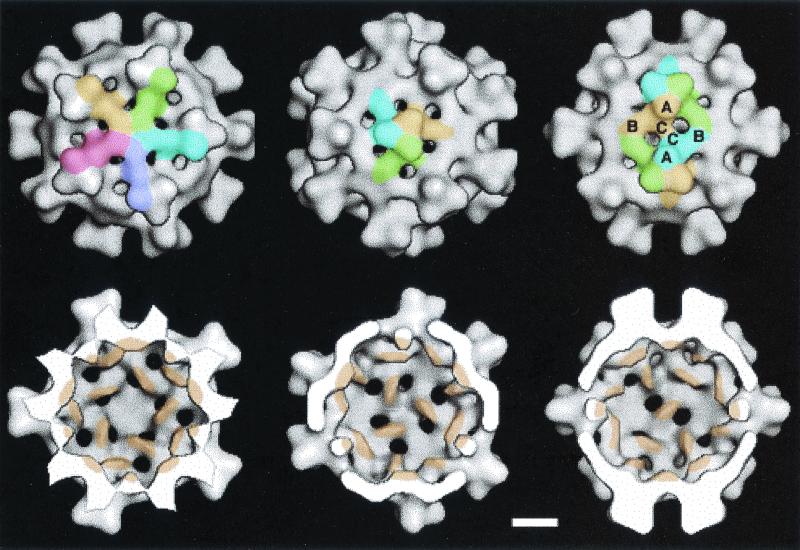
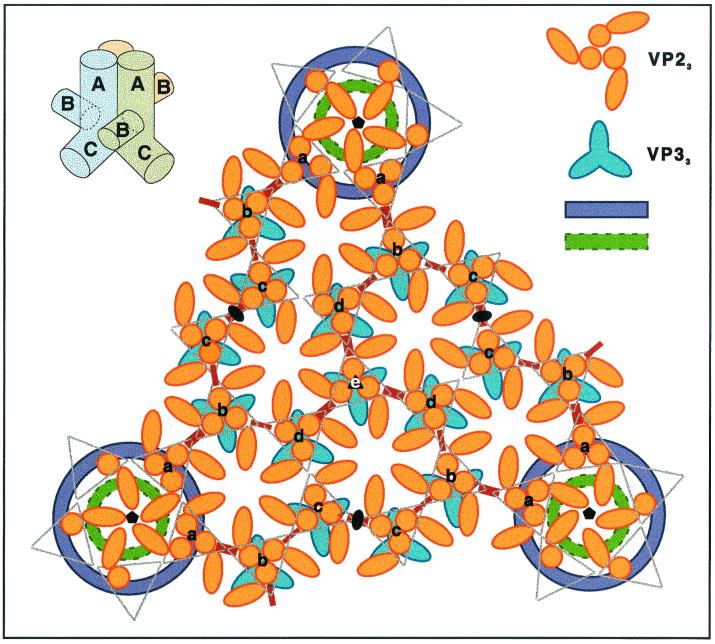
Infectious bursal disease virus (IBDV), a member of the Birnaviridae family, is a double-stranded RNA virus. The IBDV capsid is formed by two major structural proteins, VP2 and VP3, which assemble to form a T=13 markedly nonspherical capsid. During viral infection, VP2 is initially synthesized as a precursor, called VPX, whose C end is proteolytically processed to the mature form during capsid assembly. We have computed three-dimensional maps of IBDV capsid and virus-like particles built up by VP2 alone by using electron cryomicroscopy and image-processing techniques. The IBDV single-shelled capsid is characterized by the presence of 260 protruding trimers on the outer surface. Five classes of trimers can be distinguished according to their different local environments. When VP2 is expressed alone in insect cells, dodecahedral particles form spontaneously; these may be assembled into larger, fragile icosahedral capsids built up by 12 dodecahedral capsids. Each dodecahedral capsid is an empty T=1 shell composed of 20 trimeric clusters of VP2. Structural comparison between IBDV capsids and capsids consisting of VP2 alone allowed the determination of the major capsid protein locations and the interactions between them. Whereas VP2 forms the outer protruding trimers, VP3 is found as trimers on the inner surface and may be responsible for stabilizing functions. Since elimination of the C-terminal region of VPX is correlated with the assembly of T=1 capsids, this domain might be involved (either alone or in cooperation with VP3) in the induction of different conformations of VP2 during capsid morphogenesis.
Figures

References
-
- Al-Khayat H A, Bhella D, Kenney J M, Roth J-F, Kingsman A J, Martin-Rendon E, Saibil H R. Yeast Ty retrotransposons assemble into virus-like particles whose T-numbers depend on the C-terminal length of the capsid protein. J Mol Biol. 1999;292:65–73. - PubMed
-
- Azad A A, Jagadish M N, Brown M A, Hudson P J. Deletion mapping and expression in Escherichia coli of the large genomic segment of a birnavirus. Virology. 1987;161:145–152. - PubMed
-
- Baker T S, Cheng R H. A model-based approach for determining orientations of biological macromolecules imaged by cryoelectron microscopy. J Struct Biol. 1996;116:120–130. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
