

Calcineurin controls nerve activity-dependent specification of slow skeletal muscle fibers but not muscle growth
- PMID: 11606756
- PMCID: PMC60832
- DOI: 10.1073/pnas.231148598
Calcineurin controls nerve activity-dependent specification of slow skeletal muscle fibers but not muscle growth
Abstract
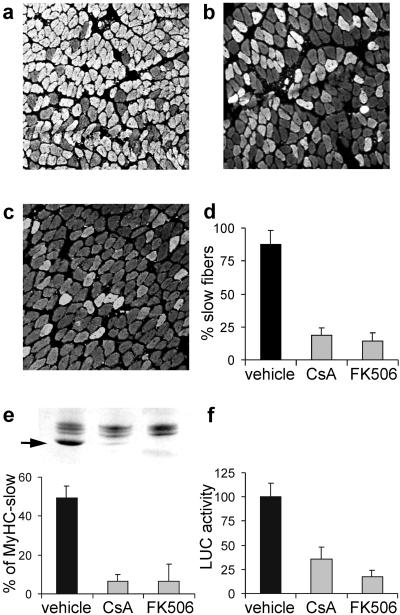
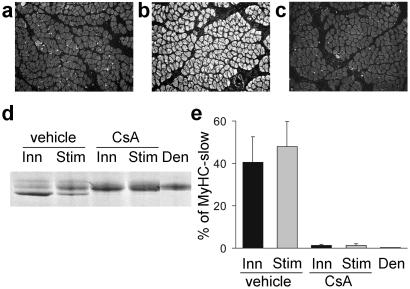
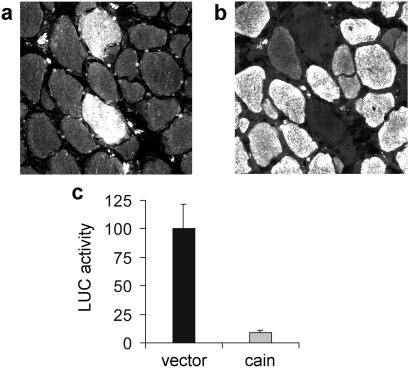
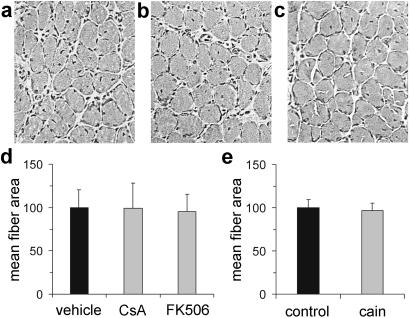
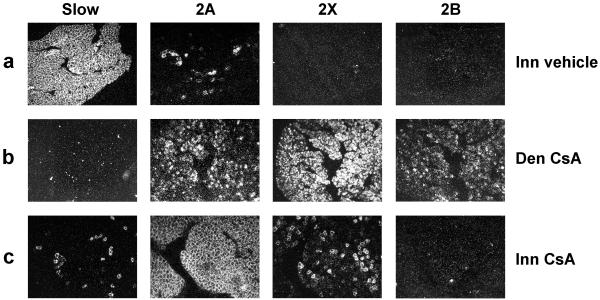
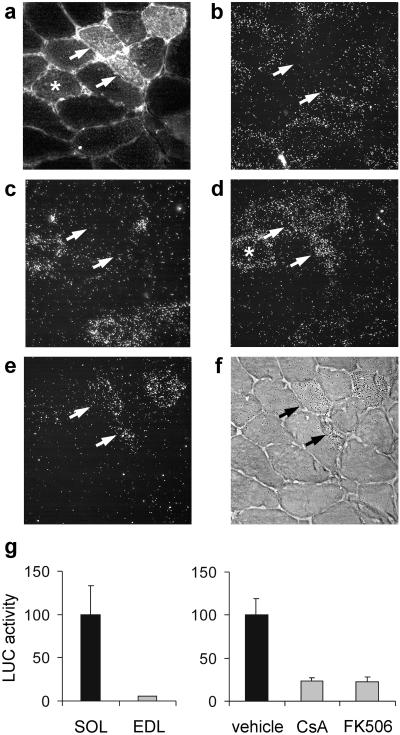
Nerve activity can induce long-lasting, transcription-dependent changes in skeletal muscle fibers and thus affect muscle growth and fiber-type specificity. Calcineurin signaling has been implicated in the transcriptional regulation of slow muscle fiber genes in culture, but the functional role of calcineurin in vivo has not been unambiguously demonstrated. Here, we report that the up-regulation of slow myosin heavy chain (MyHC) and a MyHC-slow promoter induced by slow motor neurons in regenerating rat soleus muscle is prevented by the calcineurin inhibitors cyclosporin A (CsA), FK506, and the calcineurin inhibitory protein domain from cain/cabin-1. In contrast, calcineurin inhibitors do not block the increase in fiber size induced by nerve activity in regenerating muscle. The activation of MyHC-slow induced by direct electrostimulation of denervated regenerating muscle with a continuous low frequency impulse pattern is blocked by CsA, showing that calcineurin function in muscle fibers and not in motor neurons is responsible for nerve-dependent specification of slow muscle fibers. Calcineurin is also involved in the maintenance of the slow muscle fiber gene program because in the adult soleus muscle, cain causes a switch from MyHC-slow to fast-type MyHC-2X and MyHC-2B gene expression, and the activity of the MyHC-slow promoter is inhibited by CsA and FK506.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
