

Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans
- PMID: 11641272
- PMCID: PMC312808
- DOI: 10.1101/gad.927801
Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans
Abstract
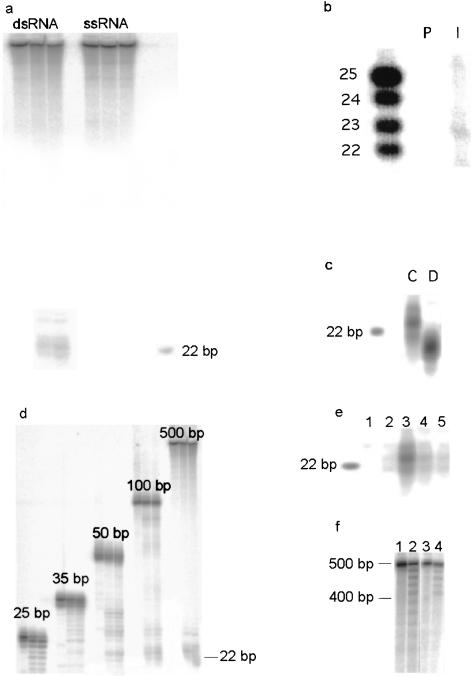
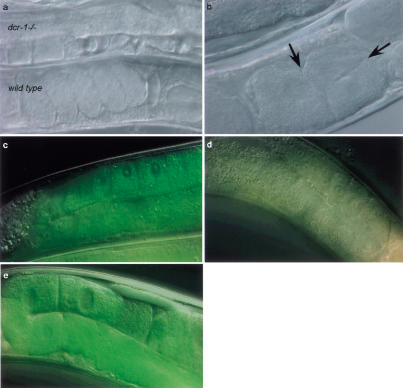
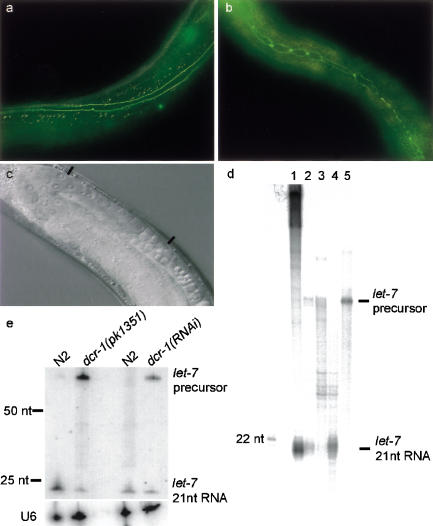
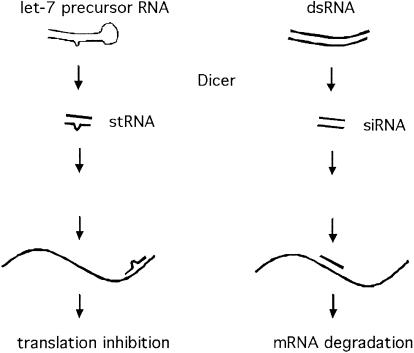
Double-stranded RNAs can suppress expression of homologous genes through an evolutionarily conserved process named RNA interference (RNAi) or post-transcriptional gene silencing (PTGS). One mechanism underlying silencing is degradation of target mRNAs by an RNP complex, which contains approximately 22 nt of siRNAs as guides to substrate selection. A bidentate nuclease called Dicer has been implicated as the protein responsible for siRNA production. Here we characterize the Caenorhabditis elegans ortholog of Dicer (K12H4.8; dcr-1) in vivo and in vitro. dcr-1 mutants show a defect in RNAi. Furthermore, a combination of phenotypic abnormalities and RNA analysis suggests a role for dcr-1 in a regulatory pathway comprised of small temporal RNA (let-7) and its target (e.g., lin-41).
Figures
References
-
- Bass BL. Double-stranded RNA as a template for gene silencing. Cell. 2000;101:235–238. - PubMed
-
- Baulcombe DC. Gene silencing: RNA makes RNA makes no protein. Curr Biol. 1999;9:R599–R601. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials