

Convergence of multiple autophagy and cytoplasm to vacuole targeting components to a perivacuolar membrane compartment prior to de novo vesicle formation
- PMID: 11675395
- PMCID: PMC2754695
- DOI: 10.1074/jbc.M109134200
Convergence of multiple autophagy and cytoplasm to vacuole targeting components to a perivacuolar membrane compartment prior to de novo vesicle formation
Abstract
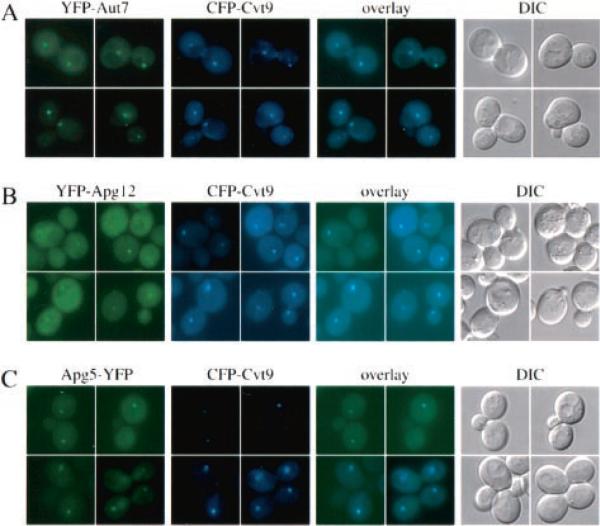
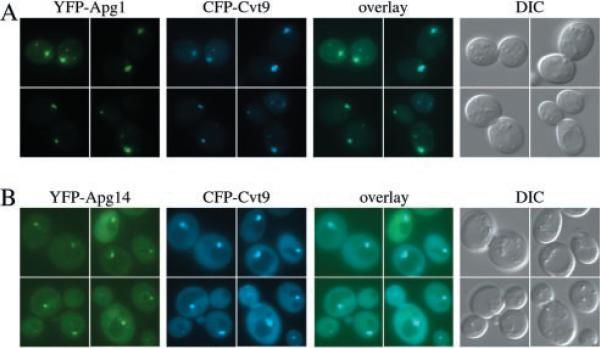
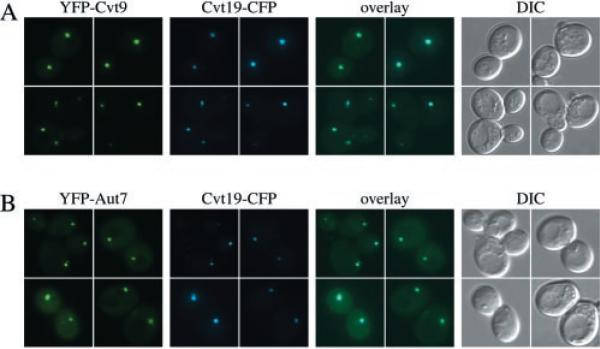
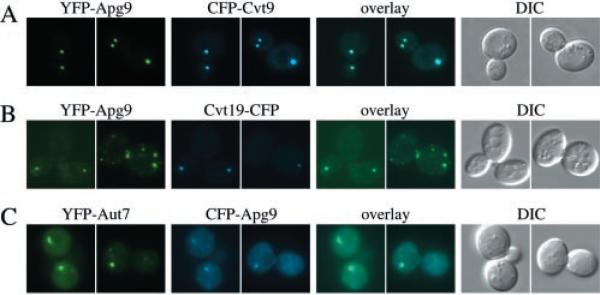
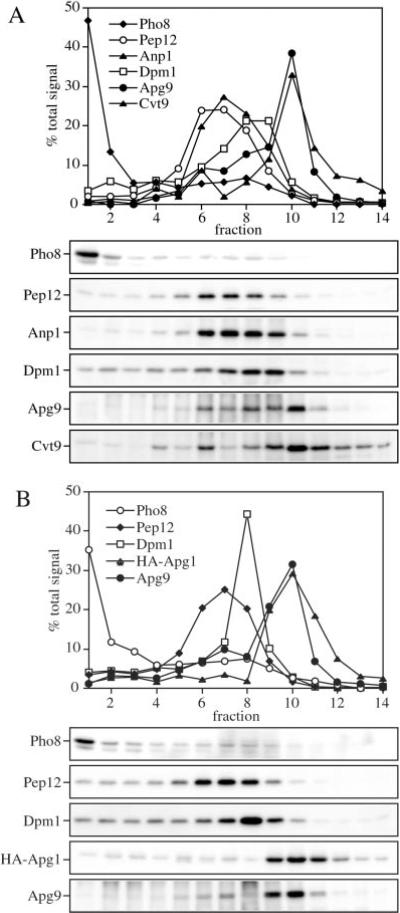
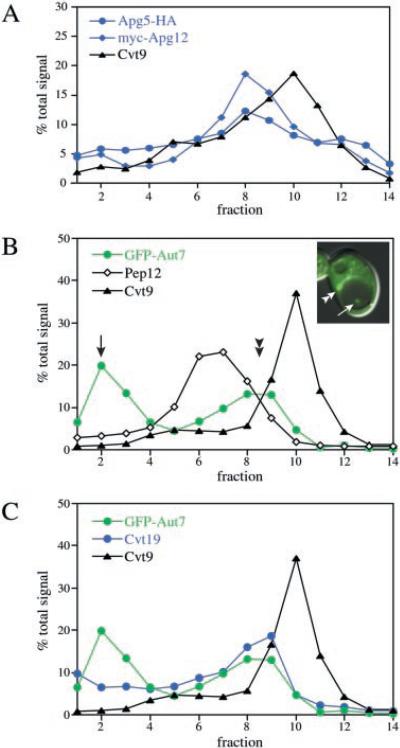
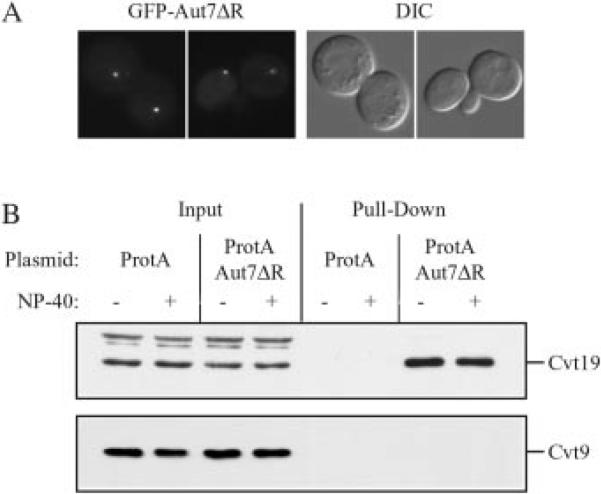
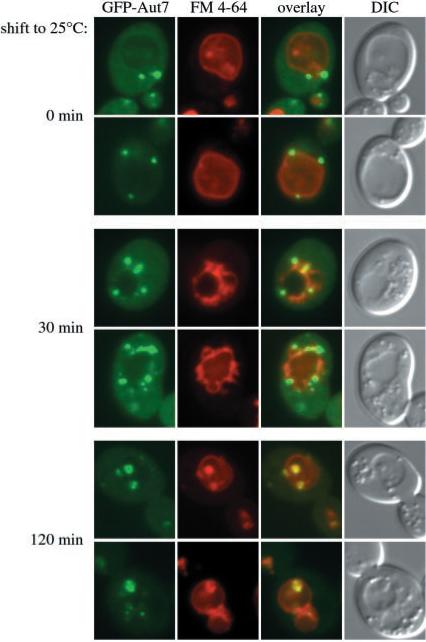
Under starvation conditions, the majority of intracellular degradation occurs at the lysosome or vacuole by the autophagy pathway. The cytoplasmic substrates destined for degradation are packaged inside unique double-membrane transport vesicles called autophagosomes and are targeted to the lysosome/vacuole for subsequent breakdown and recycling. Genetic analyses of yeast autophagy mutants, apg and aut, have begun to identify the molecular machinery as well as indicate a substantial overlap with the biosynthetic cytoplasm to vacuole targeting (Cvt) pathway. Transport vesicle formation is a key regulatory step of both pathways. In this study, we characterize the putative compartment from which both autophagosomes and the analogous Cvt vesicles may originate. Microscopy analyses identified a perivacuolar membrane as the resident compartment for both the Apg1-Cvt9 signaling complex, which mediates the switching between autophagic and Cvt transport, and the autophagy/Cvt-specific phosphatidylinositol 3-kinase complex. Furthermore, the perivacuolar compartment designates the initial site of membrane binding by the Apg/Cvt vesicle component Aut7, the Cvt cargo receptor Cvt19, and the Apg conjugation machinery, which functions in the de novo formation of vesicles. Biochemical isolation of the vesicle component Aut7 and density gradient analyses recapitulate the microscopy findings although also supporting the paradigm that components required for vesicle formation and packaging concentrate at subdomains within the donor membrane compartment.
Figures
References
-
- Paglin S, Hollister T, Delohery T, Hackett N, McMahill M, Sphicas E, Domingo D, Yahalom J. Cancer Res. 2001;61:439–444. - PubMed
-
- Bursch W, Ellinger A, Kienzl H, Török L, Pandey S, Sikorska M, Walker R, Hermann RS. Carcinogenesis. 1996;17:1595–1607. - PubMed
-
- Cavallini G, Donati A, Gori Z, Pollera M, Bergamini E. Exp. Gerontol. 2001;36:497–506. - PubMed
-
- Bursch W. Cell Death Differ. 2001;8:569–581. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
