

A biophysical model of bidirectional synaptic plasticity: dependence on AMPA and NMDA receptors
- PMID: 11675507
- PMCID: PMC60129
- DOI: 10.1073/pnas.201404598
A biophysical model of bidirectional synaptic plasticity: dependence on AMPA and NMDA receptors
Abstract
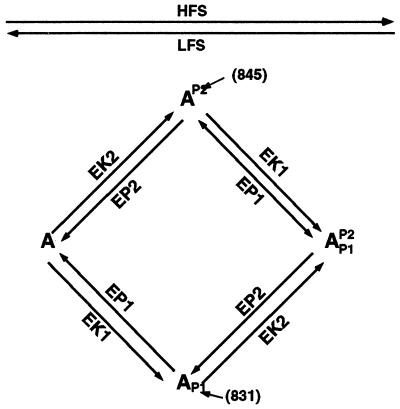
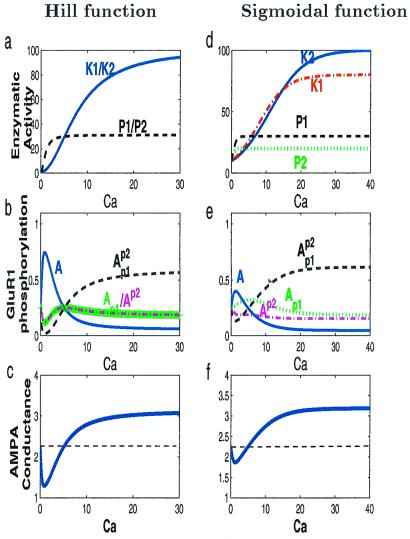
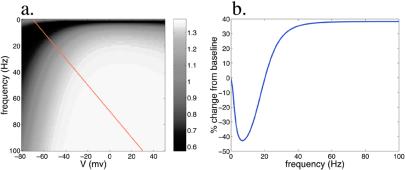
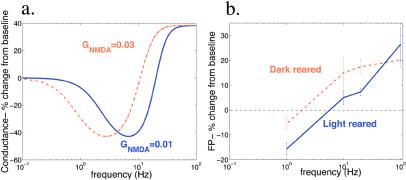
In many regions of the brain, including the mammalian cortex, the magnitude and direction of activity-dependent changes in synaptic strength depend on the frequency of presynaptic stimulation (synaptic plasticity), as well as the history of activity at those synapses (metaplasticity). We present a model of a molecular mechanism of bidirectional synaptic plasticity based on the observation that long-term synaptic potentiation (LTP) and long-term synaptic depression (LTD) correlate with the phosphorylation/dephosphorylation of sites on the alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor subunit protein GluR1. The primary assumption of the model, for which there is wide experimental support, is that postsynaptic calcium concentration and consequent activation of calcium-dependent protein kinases and phosphatases are the triggers for the induction of LTP/LTD. As calcium influx through the n-methyl-d-aspartate (NMDA) receptor plays a fundamental role in the induction of LTP/LTD, changes in the properties of NMDA receptor-mediated calcium influx will dramatically affect activity-dependent synaptic plasticity (metaplasticity). We demonstrate that experimentally observed metaplasticity can be accounted for by activity-dependent regulation of NMDA receptor subunit composition and function. Our model produces a frequency-dependent LTP/LTD curve with a sliding synaptic modification threshold similar to what has been proposed theoretically by Bienenstock, Cooper, and Munro and observed experimentally.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
