

Allergen-induced murine upper airway inflammation: local and systemic changes in murine experimental allergic rhinitis
- PMID: 11683963
- PMCID: PMC1783291
- DOI: 10.1046/j.1365-2567.2001.01253.x
Allergen-induced murine upper airway inflammation: local and systemic changes in murine experimental allergic rhinitis
Abstract
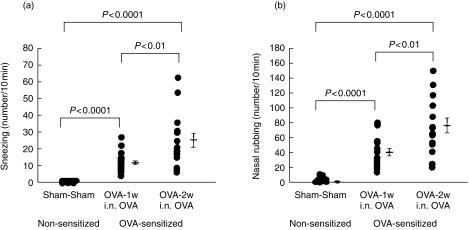
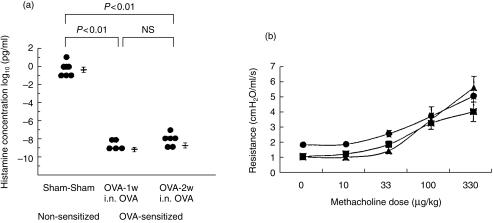
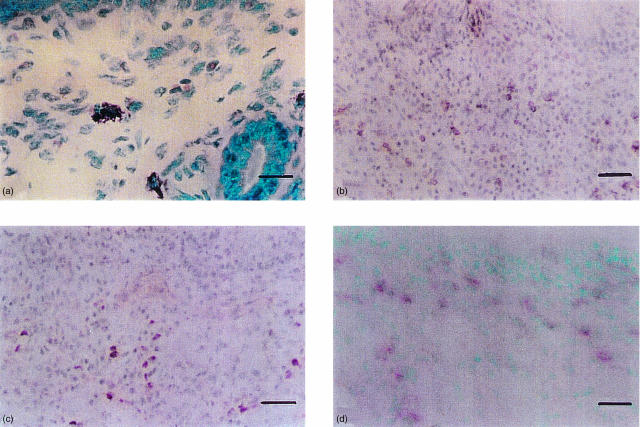
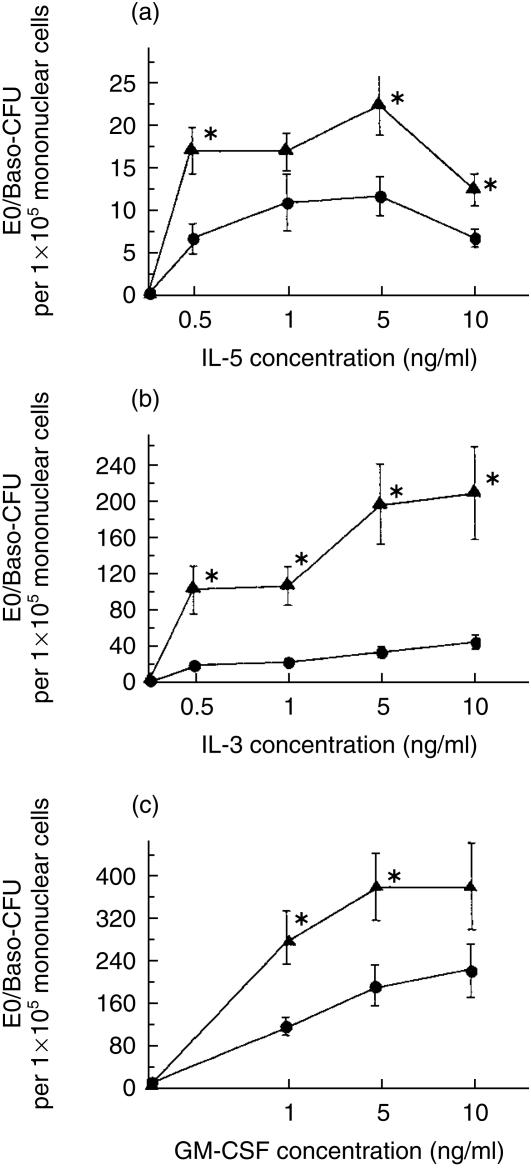
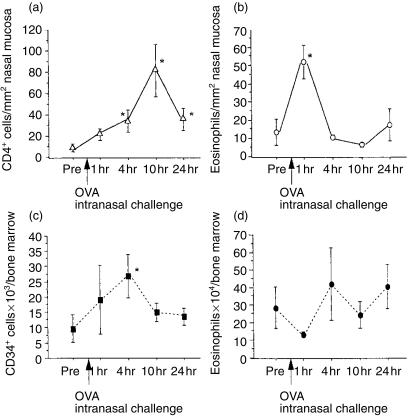
The role of inflammatory effector cells in the pathogenesis of airway allergy has been the subject of much investigation. However, whether systemic factors are involved in the development of local responses in both upper and lower airways has not been fully clarified. The present study was performed to investigate aspects of the pathogenesis of isolated allergic rhinitis in a murine model sensitized to ovalbumin (OVA). Both upper- and lower-airway physiological responsiveness and inflammatory changes were assessed, as well as bone marrow progenitor responses, by culture and immunohistological methods. Significant nasal symptoms and hyper-responsiveness appeared after intranasal OVA challenge (P < 0.0001 and P < 0.01, respectively), accompanied with significant nasal mucosal changes in CD4+ cells (P < 0.001), interleukin (IL)-4+ cells (P < 0.01), IL-5+ cells (P < 0.01), basophilic cells (P < 0.02) and eosinophils (P < 0.001), in the complete absence of hyper-responsiveness or inflammatory changes in the lower airway. In the bone marrow, there were significant increases in CD34+ cells, as well as in eosinophils and basophilic cells. In the presence in vitro of mouse recombinant IL-5, IL-3 or granulocyte-macrophage colony-stimulating factor (GM-CSF), the level of bone marrow eosinophil/basophil (Eo/Baso) colony-forming cells increased significantly in the OVA-sensitized group. We conclude that, in this murine model of allergic rhinitis, haemopoietic progenitors are upregulated, which is consistent with the involvement of bone marrow in the pathogenesis of nasal mucosal inflammation. Both local and systemic events, initiated in response to allergen provocation, may be required for the pathogenesis of allergic rhinitis. Understanding these events and their regulation could provide new therapeutic targets for rhinitis and asthma.
Figures
References
-
- Inman MD, Ellis R, Wattie J, Denburg JA, O'Byrne PM. Allergen-induced increase in airway responsiveness, airway eosinophilia and bone-marrow eosinophil progenitors in mice. Am J Respir Cell Mol Biol. 1999;21:473–9. PM. - PubMed
-
- Gaspar Elsas MI, Joseph D, Elsas PX, Vargaftig BB. Rapid increase in bone marrow eosinphil production and responses to eosinopoietic interleukins triggered by intranasal allergen challenge. Am J Respir Cell Mol Biol. 1997;17:404–13. - PubMed
-
- Denburg JA, Telizyn S, Belda A, Dolovich J, Bienenstock J. Increased numbers of circulating basophil progenitors in atopic patients. J Allergy Clin Immunol. 1985;76:466–72. - PubMed
-
- Otsuka H, Dolovich J, Befus AD, Telizyn S, Bienenstock J, Denburg JA. Basophilic cell progenitors, nasal metachromatic cells, and peripheral blood basophils in ragweed-allergic patients. J Allergy Clin Immunol. 1986;78:365–71. - PubMed
-
- Gibson PG, Dolovich J, Girgis-Gabardo A, Morris MM, Anderson M, Hargreave FE, Denburg JA. The inflammatory response in asthma exacerbation: changes in circulating eosinophils, basophils and their progenitors. Clin Exp Allergy. 1990;20:661–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
