

Intraflagellar transport balances continuous turnover of outer doublet microtubules: implications for flagellar length control
- PMID: 11684707
- PMCID: PMC2150833
- DOI: 10.1083/jcb.200106141
Intraflagellar transport balances continuous turnover of outer doublet microtubules: implications for flagellar length control
Abstract
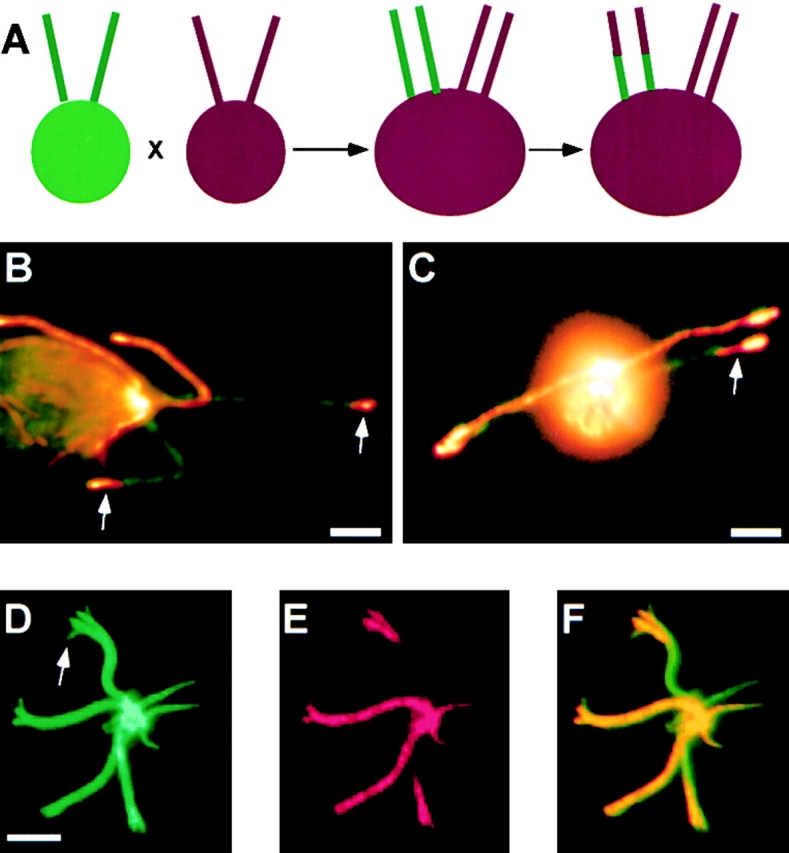
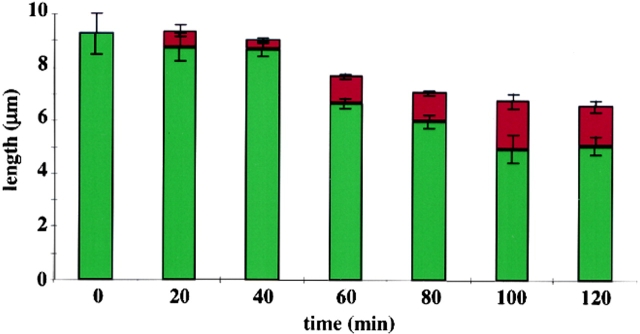
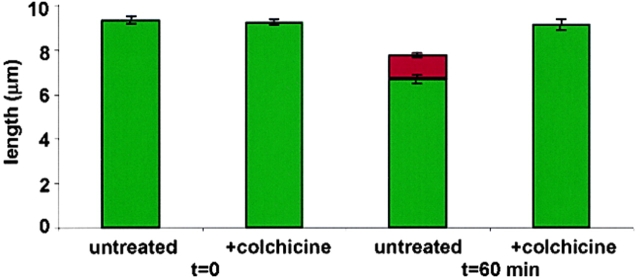
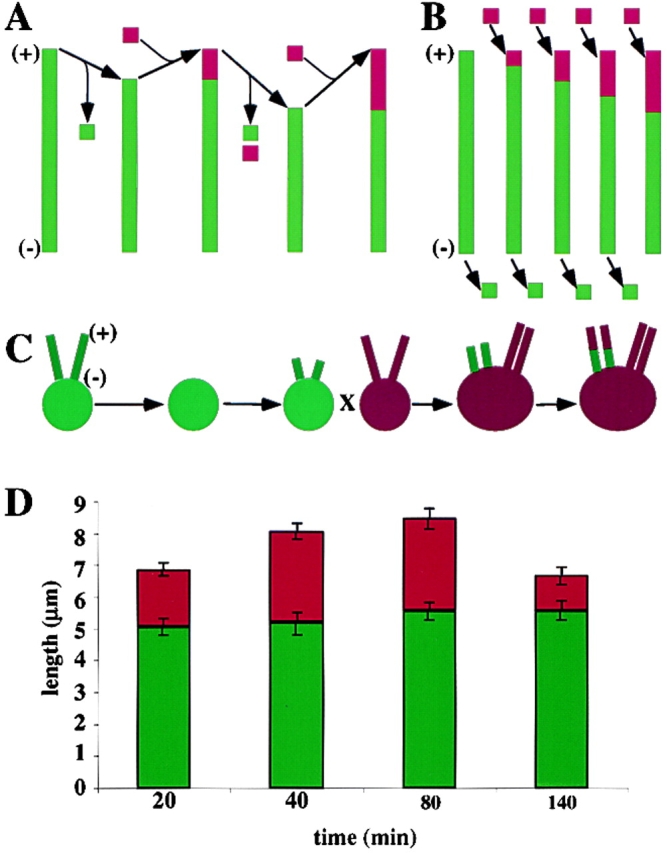
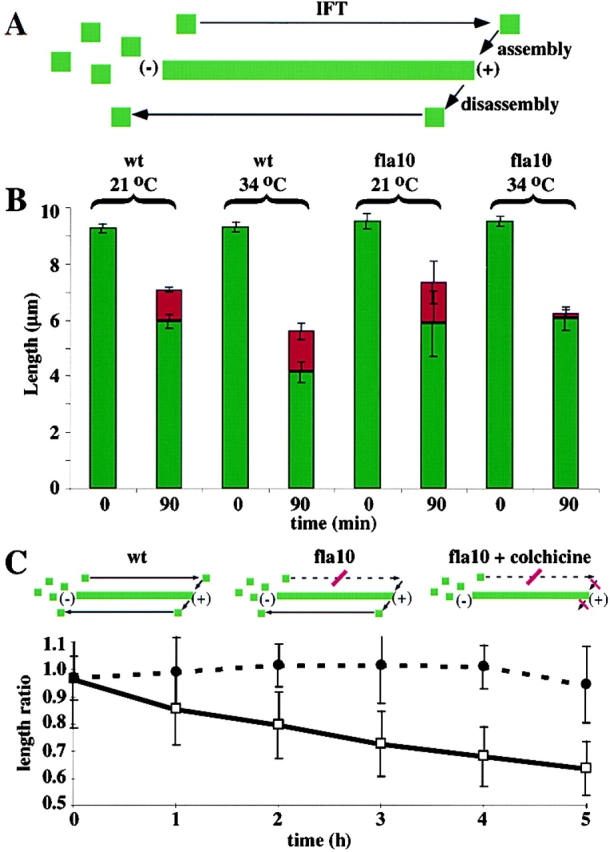
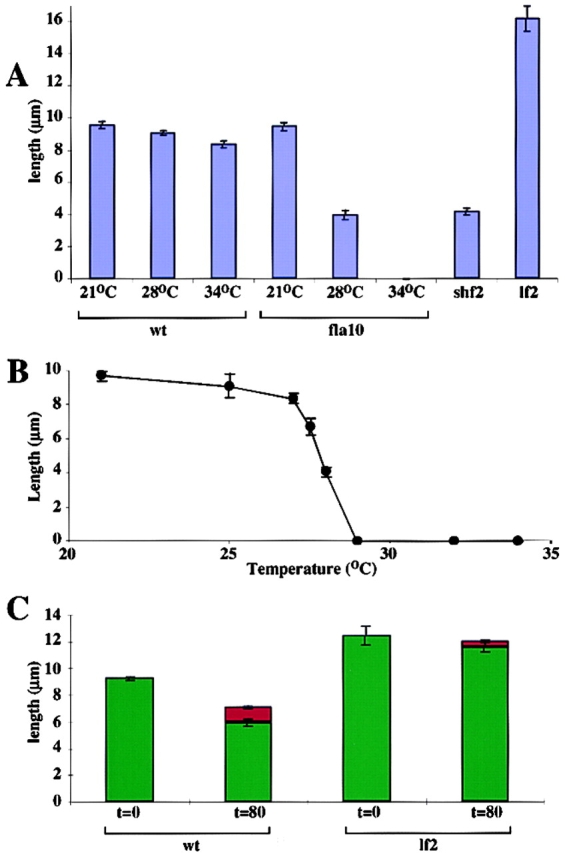
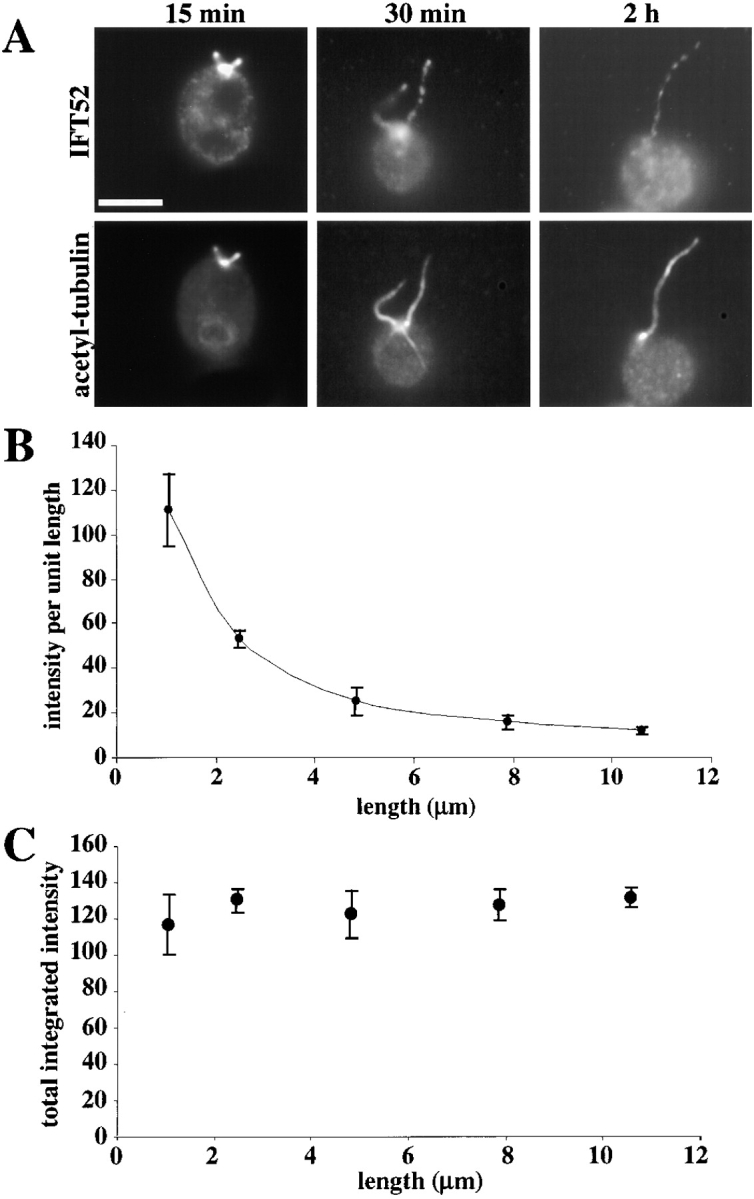
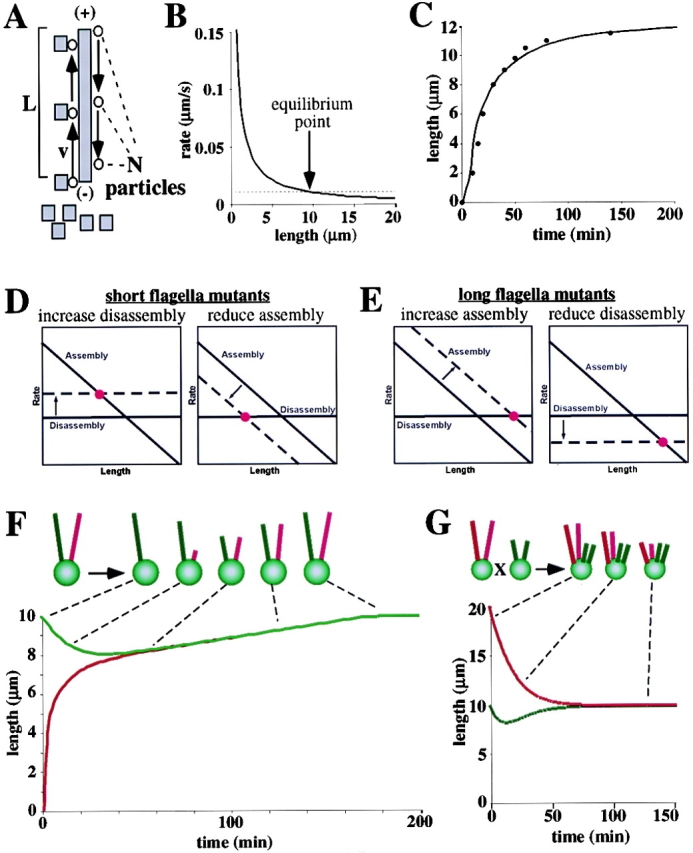
A central question in cell biology is how cells determine the size of their organelles. Flagellar length control is a convenient system for studying organelle size regulation. Mechanistic models proposed for flagellar length regulation have been constrained by the assumption that flagella are static structures once they are assembled. However, recent work has shown that flagella are dynamic and are constantly turning over. We have determined that this turnover occurs at the flagellar tips, and that the assembly portion of the turnover is mediated by intraflagellar transport (IFT). Blocking IFT inhibits the incorporation of tubulin at the flagellar tips and causes the flagella to resorb. These results lead to a simple steady-state model for flagellar length regulation by which a balance of assembly and disassembly can effectively regulate flagellar length.
Figures
References
-
- Behnke, O., and A. Forer. 1967. Evidence for four classes of microtubules in individual cells. J. Cell Sci. 2:169–192. - PubMed
-
- Cole, D.G., D.R. Diener, A.L. Himelblau, P.L. Beech, J.C. Fuster, and J.L. Rosenbaum. 1998. Chlamydomonas kinesin-II–dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis elegans sensory cilia. J. Cell Biol. 141:993–1008. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
