

How matrix metalloproteinases regulate cell behavior
- PMID: 11687497
- PMCID: PMC2792593
- DOI: 10.1146/annurev.cellbio.17.1.463
How matrix metalloproteinases regulate cell behavior
Abstract
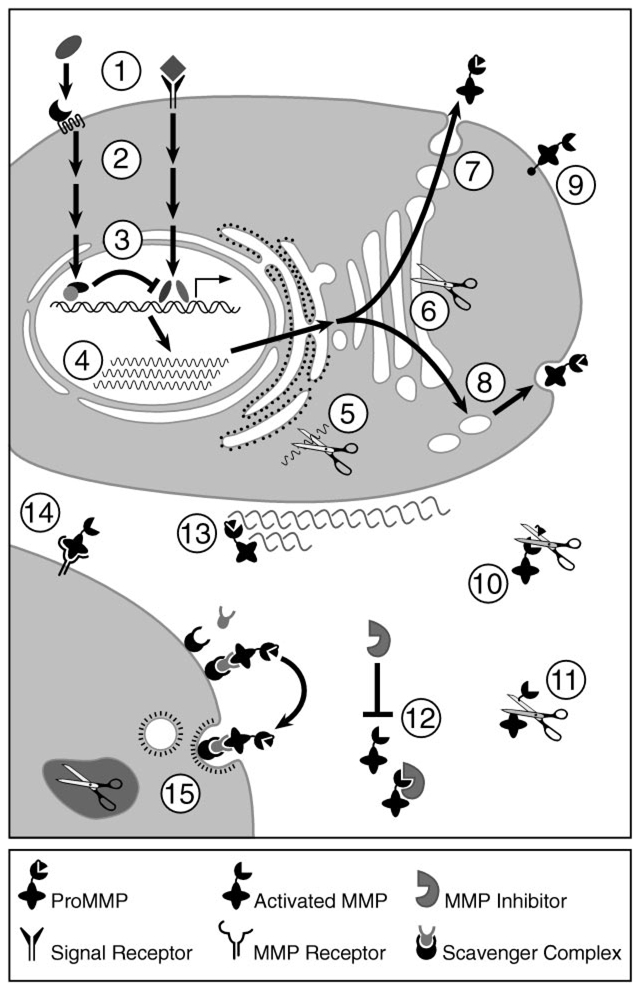
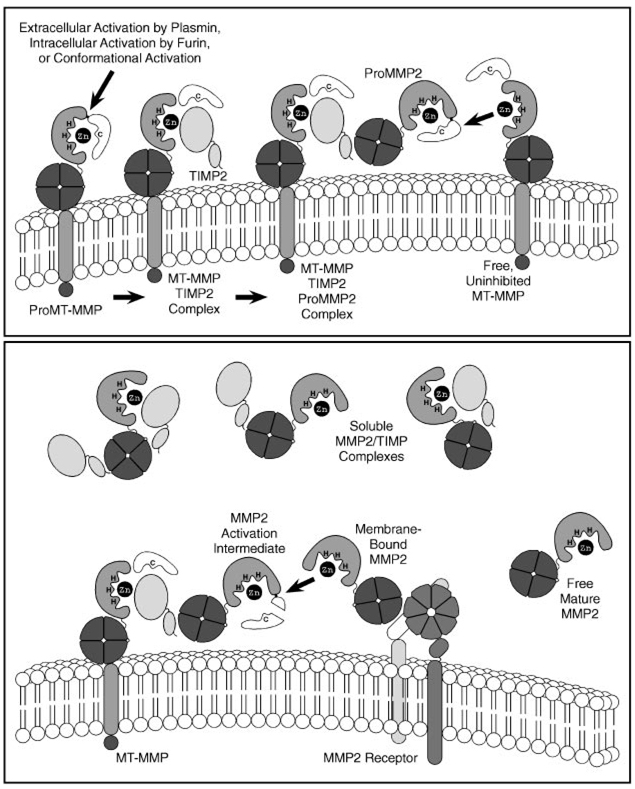
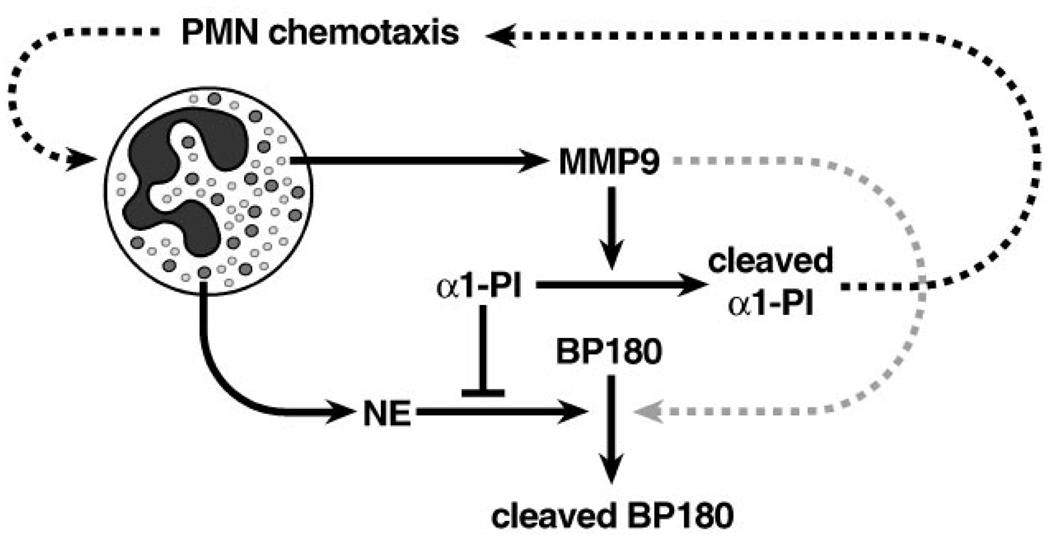
The matrix metalloproteinases (MMPs) constitute a multigene family of over 25 secreted and cell surface enzymes that process or degrade numerous pericellular substrates. Their targets include other proteinases, proteinase inhibitors, clotting factors, chemotactic molecules, latent growth factors, growth factor-binding proteins, cell surface receptors, cell-cell adhesion molecules, and virtually all structural extracellular matrix proteins. Thus MMPs are able to regulate many biologic processes and are closely regulated themselves. We review recent advances that help to explain how MMPs work, how they are controlled, and how they influence biologic behavior. These advances shed light on how the structure and function of the MMPs are related and on how their transcription, secretion, activation, inhibition, localization, and clearance are controlled. MMPs participate in numerous normal and abnormal processes, and there are new insights into the key substrates and mechanisms responsible for regulating some of these processes in vivo. Our knowledge in the field of MMP biology is rapidly expanding, yet we still do not fully understand how these enzymes regulate most processes of development, homeostasis, and disease.
Figures


References
-
- Abbaszade I, Liu RQ, Yang F, Rosenfeld SA, Ross OH, et al. Cloning and characterization of ADAMTS11, an aggrecanase from the ADAMTS family. J. Biol. Chem. 1999;274:23443–23450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
