

Increased levels of keratin 16 alter epithelialization potential of mouse skin keratinocytes in vivo and ex vivo
- PMID: 11694579
- PMCID: PMC60266
- DOI: 10.1091/mbc.12.11.3439
Increased levels of keratin 16 alter epithelialization potential of mouse skin keratinocytes in vivo and ex vivo
Abstract
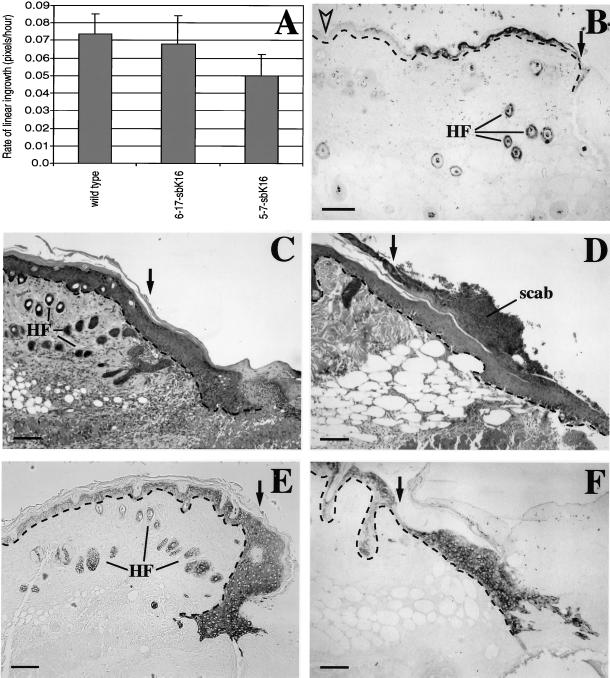
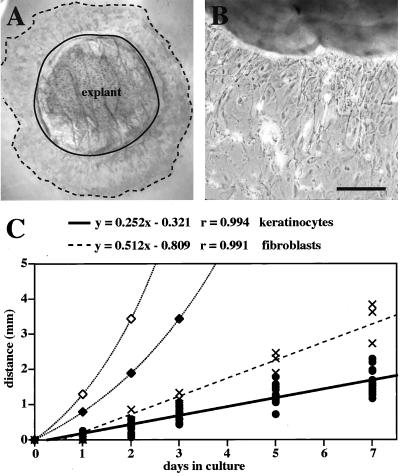
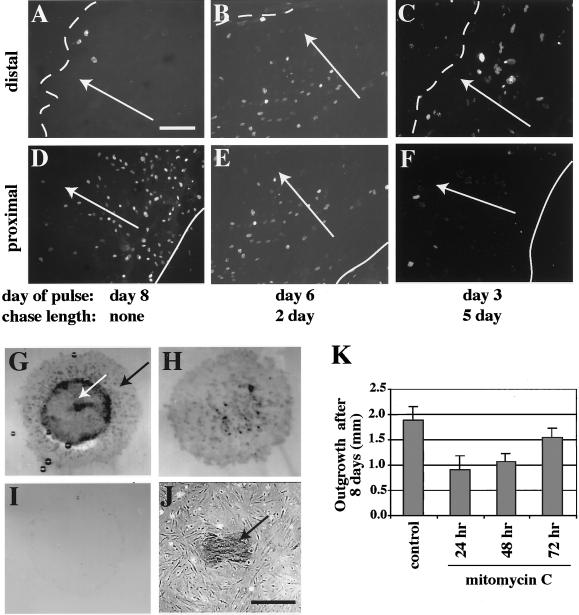
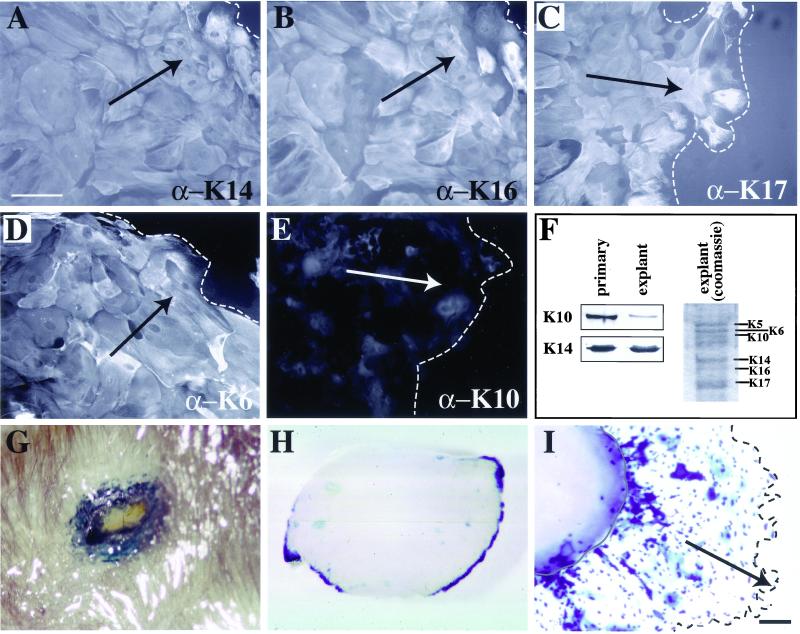
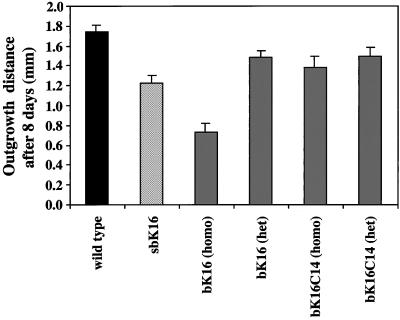
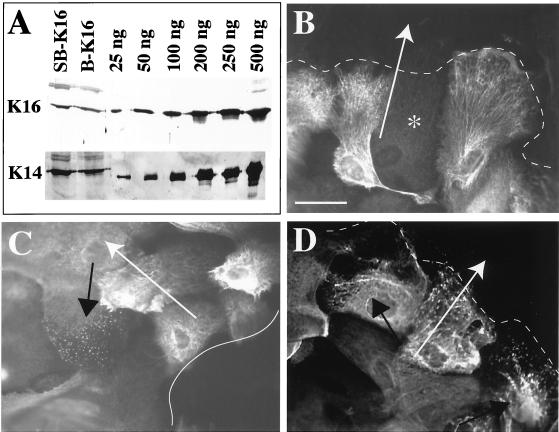
The process of wound repair in adult skin is complex, involving dermal contraction and epithelial migration to repair the lesion and restore the skin's barrier properties. At the wound edge, keratinocytes undergo many changes that engender an epithelialization behavior. The type II keratin 6 and type I keratins 16 and 17 are induced well before cell migration begins, but the role of these proteins is not understood. Forced expression of human K16 in skin epithelia of transgenic mice has been shown to cause dose-dependent skin lesions concomitant with alterations in keratin filament organization and in cell adhesion. Here we show, with the use of a quantitative assay, that these transgenic mice show a delay in the closure of full-thickness skin wounds in situ compared with wild-type and low-expressing K16 transgenic mice. We adapted and validated an ex vivo skin explant culture system to better assess epithelialization in a wound-like environment. Transgenic K16 explants exhibit a significant reduction of keratinocyte outgrowth in this setting. This delay is transgene dose-dependent, and is more severe when K16 is expressed in mitotic compared with post-mitotic keratinocytes. Various lines of evidence suggest that the mechanism(s) involved is complex and not strictly cell autonomous. These findings have important implications for the function of K16 in vivo.
Figures
Similar articles
-
Forced expression of keratin 16 alters the adhesion, differentiation, and migration of mouse skin keratinocytes.Mol Biol Cell. 2000 Oct;11(10):3315-27. doi: 10.1091/mbc.11.10.3315. Mol Biol Cell. 2000. PMID: 11029038 Free PMC article.
-
Onset of re-epithelialization after skin injury correlates with a reorganization of keratin filaments in wound edge keratinocytes: defining a potential role for keratin 16.J Cell Biol. 1996 Feb;132(3):381-97. doi: 10.1083/jcb.132.3.381. J Cell Biol. 1996. PMID: 8636216 Free PMC article.
-
An ex vivo assay to assess the potential of skin keratinocytes for wound epithelialization.J Invest Dermatol. 2002 May;118(5):866-70. doi: 10.1046/j.1523-1747.2002.01736.x. J Invest Dermatol. 2002. PMID: 11982766
-
Overexpression of human keratin 16 produces a distinct skin phenotype in transgenic mouse skin.Biochem Cell Biol. 1995 Sep-Oct;73(9-10):611-8. doi: 10.1139/o95-067. Biochem Cell Biol. 1995. PMID: 8714680 Review.
-
Re-epithelialization. Human keratinocyte locomotion.Dermatol Clin. 1993 Oct;11(4):641-6. Dermatol Clin. 1993. PMID: 8222348 Review.
Cited by
-
The skin transcriptome in hidradenitis suppurativa uncovers an antimicrobial and sweat gland gene signature which has distinct overlap with wounded skin.PLoS One. 2019 May 6;14(5):e0216249. doi: 10.1371/journal.pone.0216249. eCollection 2019. PLoS One. 2019. PMID: 31059533 Free PMC article.
-
Exploring a Role for Regulatory miRNAs In Wound Healing during Ageing:Involvement of miR-200c in wound repair.Sci Rep. 2017 Jun 12;7(1):3257. doi: 10.1038/s41598-017-03331-6. Sci Rep. 2017. PMID: 28607463 Free PMC article.
-
Development of a three dimensional multiscale computational model of the human epidermis.PLoS One. 2010 Jan 14;5(1):e8511. doi: 10.1371/journal.pone.0008511. PLoS One. 2010. PMID: 20076760 Free PMC article.
-
N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis.Mar Drugs. 2019 Sep 4;17(9):520. doi: 10.3390/md17090520. Mar Drugs. 2019. PMID: 31487895 Free PMC article.
-
Vimentin is necessary for colony growth of human diploid keratinocytes.Histochem Cell Biol. 2015 Jan;143(1):45-57. doi: 10.1007/s00418-014-1262-6. Epub 2014 Aug 21. Histochem Cell Biol. 2015. PMID: 25142512
References
-
- Bereiter-Hahn J. Epidermal cell migration and wound repair. In: Bereiter-Hahn J, Matoltsy AG, Richards KS, editors. Biology of the Integument. 2, Vertebrates. Berlin: Springer-Verlag; 1986. pp. 443–471.
-
- Brock J, McCluskey J, Baribault H, Martin P. Perfect wound healing in the keratin 8 deficient mouse embryo. Cell Motil Cytoskeleton. 1996;35:358–366. - PubMed
-
- Byrne C, Tainsky M, Fuchs E. Programming gene expression in developing epidermis. Development. 1994;120:2369–2383. - PubMed
-
- Chen JD, Kim JP, Zhang K, Sarret Y, Wynn KC, Kramer RH, Woodley DT. Epidermal growth factor (EGF) promotes keratinocyte locomotion on collagen by increasing the α2 integrin subunit. Exp Cell Res. 1993;209:216–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
