

Tumor necrosis factor-alpha induces stress fiber formation through ceramide production: role of sphingosine kinase
- PMID: 11694593
- PMCID: PMC60280
- DOI: 10.1091/mbc.12.11.3618
Tumor necrosis factor-alpha induces stress fiber formation through ceramide production: role of sphingosine kinase
Abstract
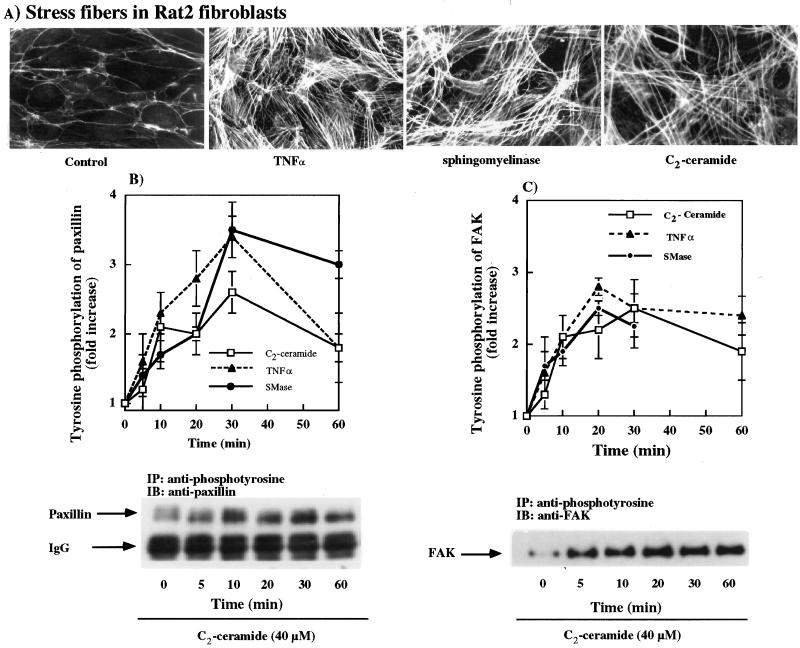
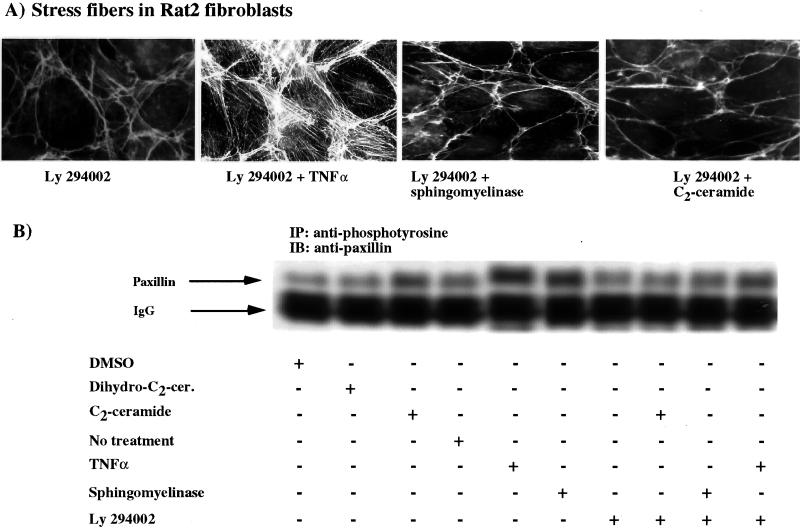
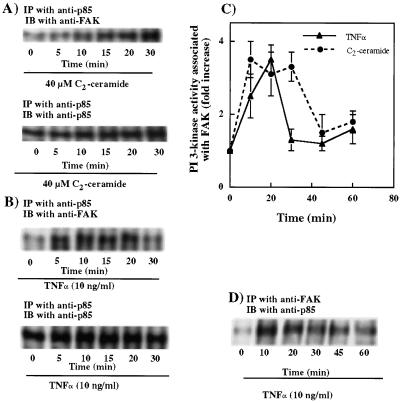
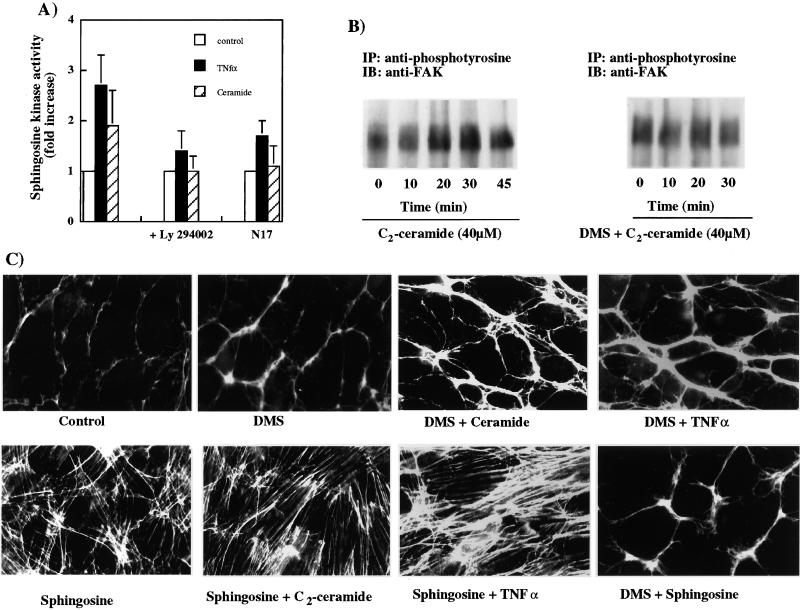
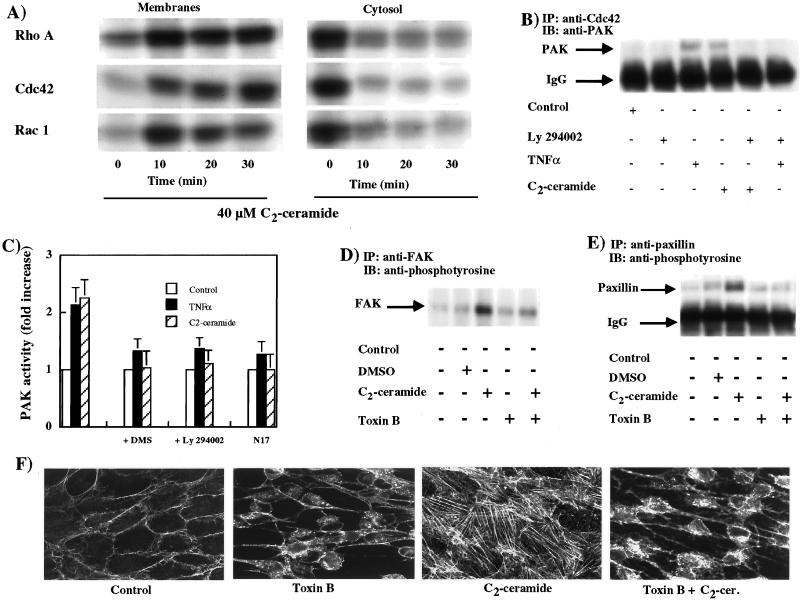
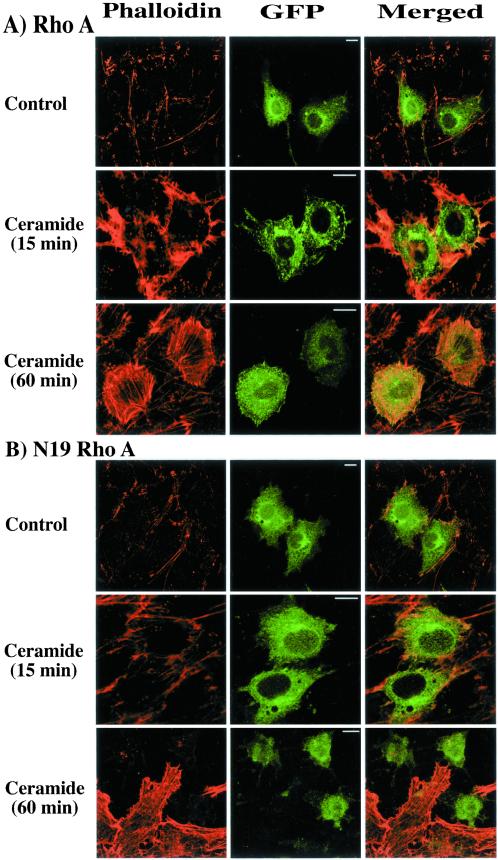
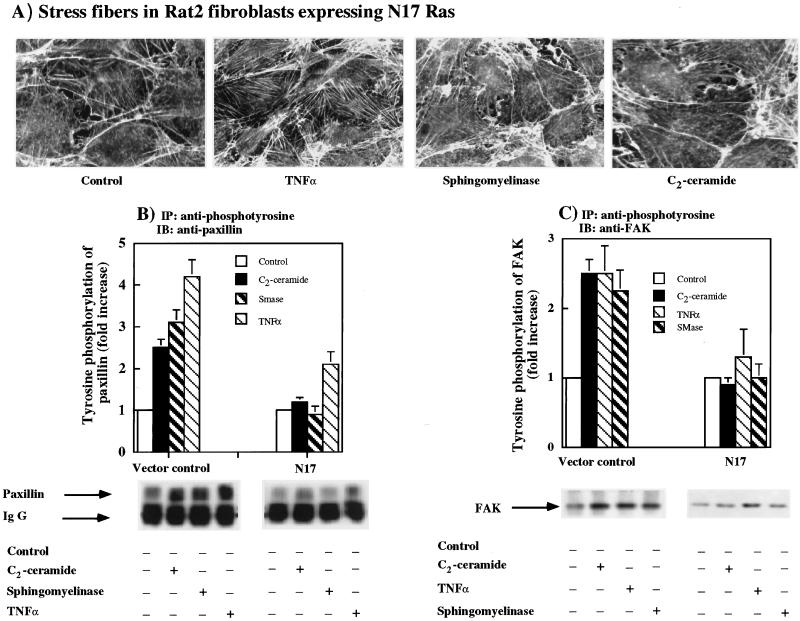
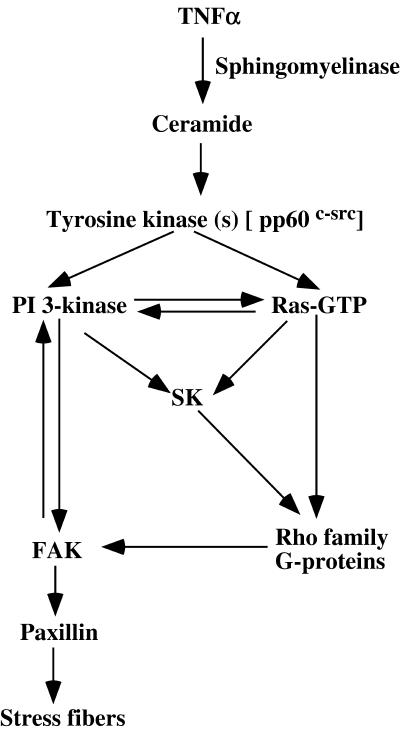
Tumor necrosis factor-alpha (TNF-alpha) is a proinflammatory cytokine that activates several signaling cascades. We determined the extent to which ceramide is a second messenger for TNF-alpha-induced signaling leading to cytoskeletal rearrangement in Rat2 fibroblasts. TNF-alpha, sphingomyelinase, or C(2)-ceramide induced tyrosine phosphorylation of focal adhesion kinase (FAK) and paxillin, and stress fiber formation. Ly 294002, a phosphatidylinositol 3-kinase (PI 3-K) inhibitor, or expression of dominant/negative Ras (N17) completely blocked C(2)-ceramide- and sphingomyelinase-induced tyrosine phosphorylation of FAK and paxillin and severely decreased stress fiber formation. The TNF-alpha effects were only partially inhibited. Dimethylsphingosine, a sphingosine kinase (SK) inhibitor, blocked stress fiber formation by TNF-alpha and C(2)-ceramide. TNF-alpha, sphingomyelinase, and C(2)-ceramide translocated Cdc42, Rac, and RhoA to membranes, and stimulated p21-activated protein kinase downstream of Ras-GTP, PI 3-K, and SK. Transfection with inactive RhoA inhibited the TNF-alpha- and C(2)-ceramide-induced stress fiber formation. Our results demonstrate that stimulation by TNF-alpha, which increases sphingomyelinase activity and ceramide formation, activates sphingosine kinase, Rho family GTPases, focal adhesion kinase, and paxillin. This novel pathway of ceramide signaling can account for approximately 70% of TNF-alpha-induced stress fiber formation and cytoskeletal reorganization.
Figures
References
-
- Abousalham A, Liossis C, O'Brien L, Brindley DN. Cell-permeable ceramides prevent the activation of phospholipase D by ADP-ribosylation factor and RhoA. J Biol Chem. 1997;272:1069–1075. - PubMed
-
- Albouz S, Boutry JM, Dubois G, Bourdon R, Hauw JJ, Baumann N. Lipid and lysosomal enzymes in human fibroblasts cultured with perhexiline maleate. Naunyn-Schmiedeberg's Arch Pharmacol. 1981;317:173–177. - PubMed
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Chant J, Stowers L. GTPase cascades choreographing cellular behavior: movement, morphogenesis, and more. Cell. 1995;81:1–4. - PubMed
-
- Chen HC, Appeddu PA, Isodo H, Guan JL. Phosphorylation of tyrosine 397 in focal adhesion kinase is required for binding phosphatidylinositol 3-kinase. J Biol Chem. 1996;271:26329–26334. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
