

Beta1 integrins show specific association with CD98 protein in low density membranes
- PMID: 11696247
- PMCID: PMC59658
- DOI: 10.1186/1471-2091-2-10
Beta1 integrins show specific association with CD98 protein in low density membranes
Abstract
Background: The CD98 (4F2, FRP-1) is a widely expressed cell surface protein heterodimer composed of a glycosylated heavy chain and a non-glycosylated light chain. Originally described as a T cell activation antigen, it was later shown to function in amino acid transport, cell fusion and homotypic cell aggregation. Several lines of evidence suggest its functional interaction with integrins but the biochemical basis for this interaction has been unclear.
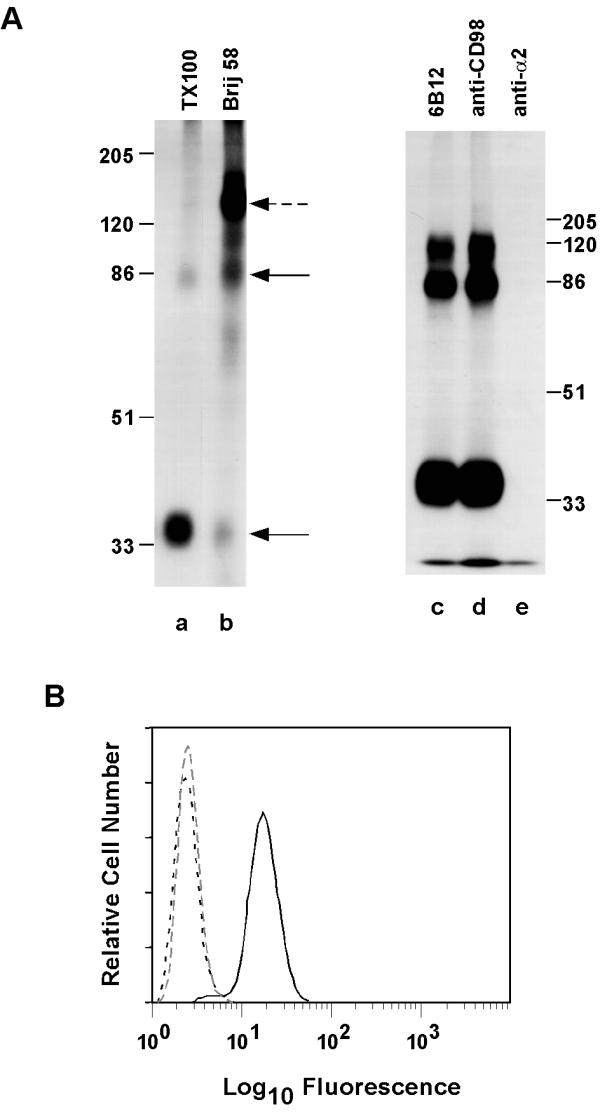
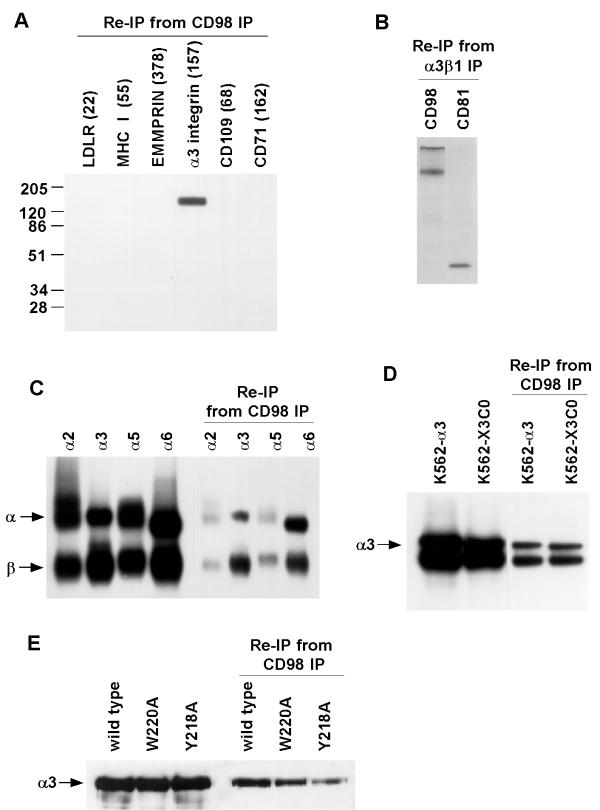
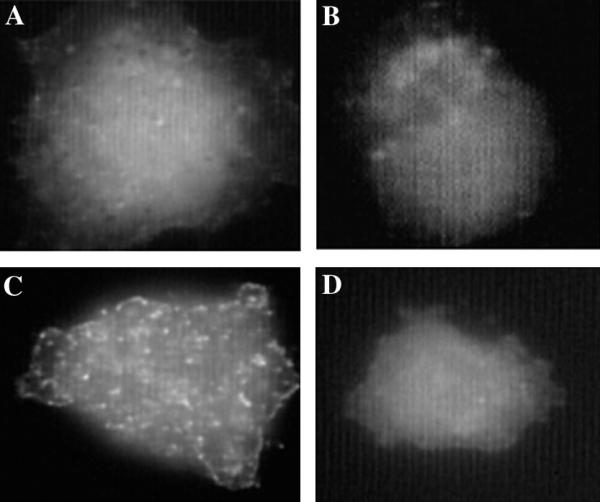
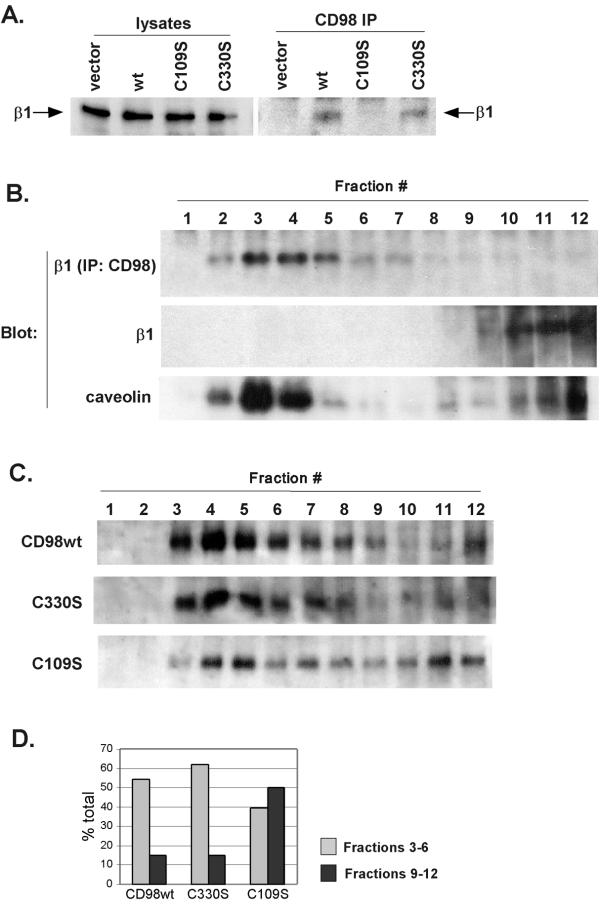
Results: We demonstrate that CD98 constitutively and specifically associates with beta1 integrins (alpha2beta1,alpha3beta1, alpha5beta1 and alpha6beta1), but minimally with alpha4beta1. Integrin-CD98 association was established by reciprocal immunoprecipitation experiments, and confirmed by CD98-induced clustering of alpha3beta1 but not alpha4beta1 on the surface of rhabdomyosarcoma cells. Integrin-CD98 association is independent of the alpha subunit cytoplasmic tail, is maintained in alpha3beta1 ligand-interaction deficient mutants, and is not inhibited by EDTA. Within the CD98 heavy chain, a C109S mutation (but not a C330S mutation) caused a loss of beta1 integrin association. The same C109S mutation also caused a loss of CD98 light chain association. Importantly, CD98 associated selectively with beta1 integrins present in low density "light membrane" fractions on a sucrose gradient. CD98 was not present in dense fractions that contained the majority of beta1 integrins. Notably, the C109S mutant of CD98, that did not associate with beta1 integrins, showed also a reduced localization into light membrane fractions.
Conclusions: We demonstrate that CD98 association with beta1 integrins is specific, occurs in the context of low density membranes, and may require the CD98 light chain.
Figures
References
-
- Haynes BF, Hemler ME, Mann DL, Eisenbarth GS, Shelhamer J, Mostowski HS, et al. Characterization of a monoclonal antibody (4F2) that binds to human monocytes and to a subset of activated lymphocytes. J Immunol. 1981;126:1409–1414. - PubMed
-
- Tabata N, Ito M, Shimokata K, Suga S, Ohgimoto S, Tsurudome M, et al. Expression of fusion regulatory proteins (FRPs) on human peripheral blood monocytes. J Immunol. 1994;153:3256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
