

alpha-Toxin is a mediator of Staphylococcus aureus-induced cell death and activates caspases via the intrinsic death pathway independently of death receptor signaling
- PMID: 11696559
- PMCID: PMC2198876
- DOI: 10.1083/jcb.200105081
alpha-Toxin is a mediator of Staphylococcus aureus-induced cell death and activates caspases via the intrinsic death pathway independently of death receptor signaling
Abstract
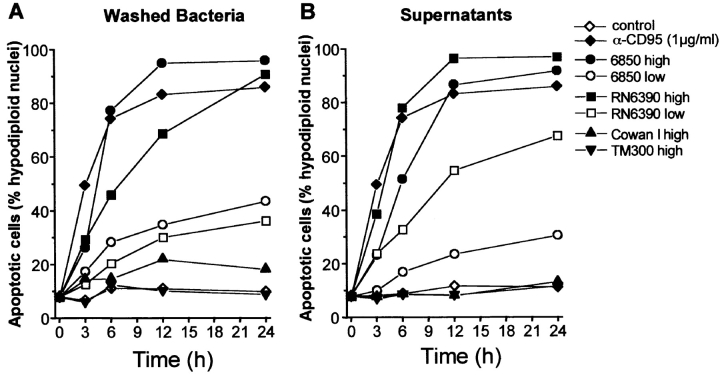
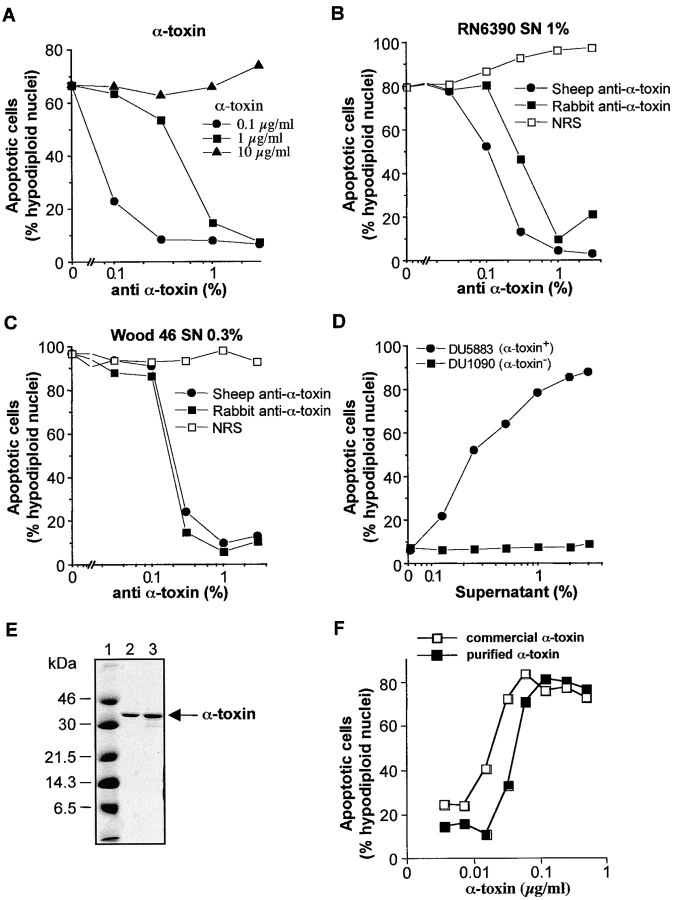
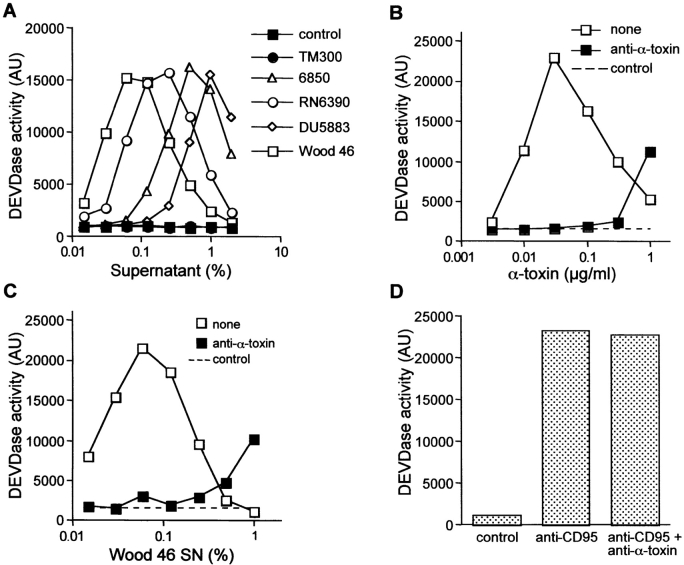
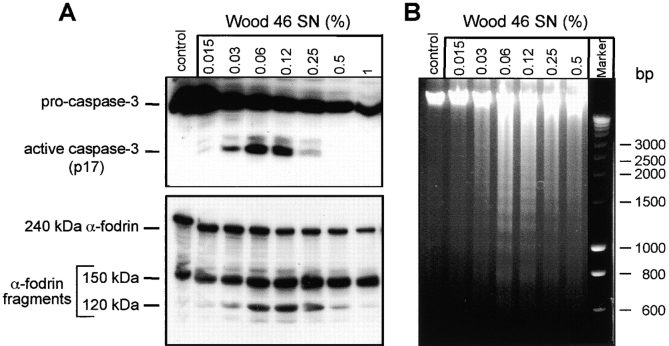
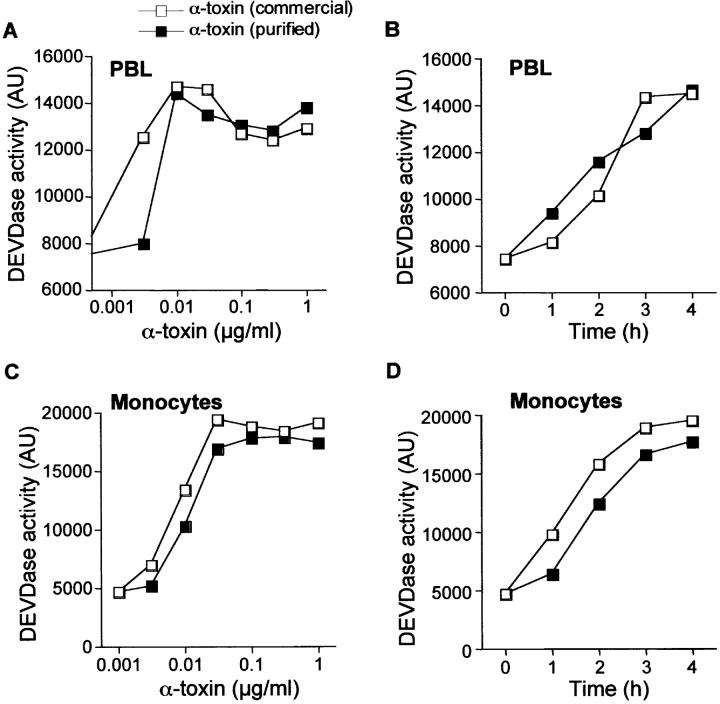
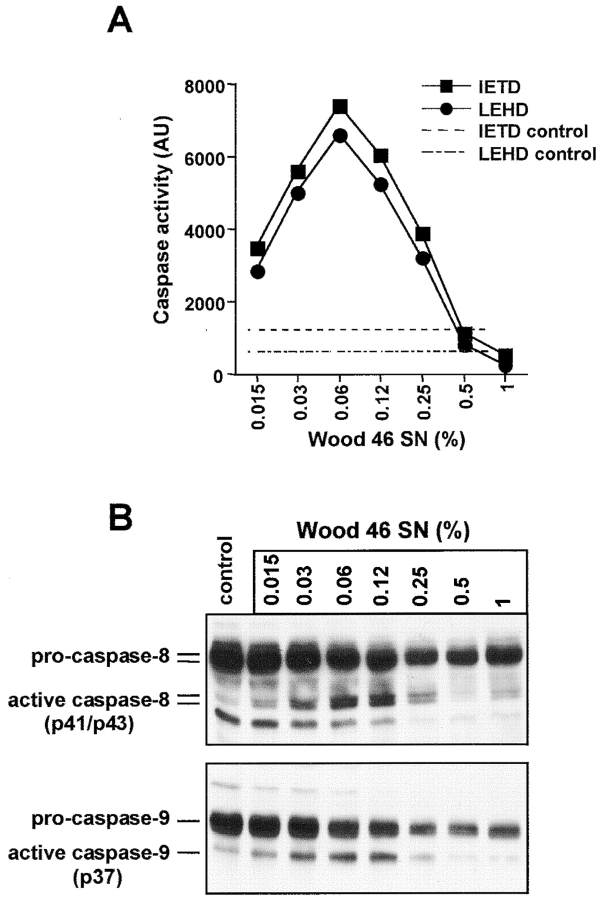
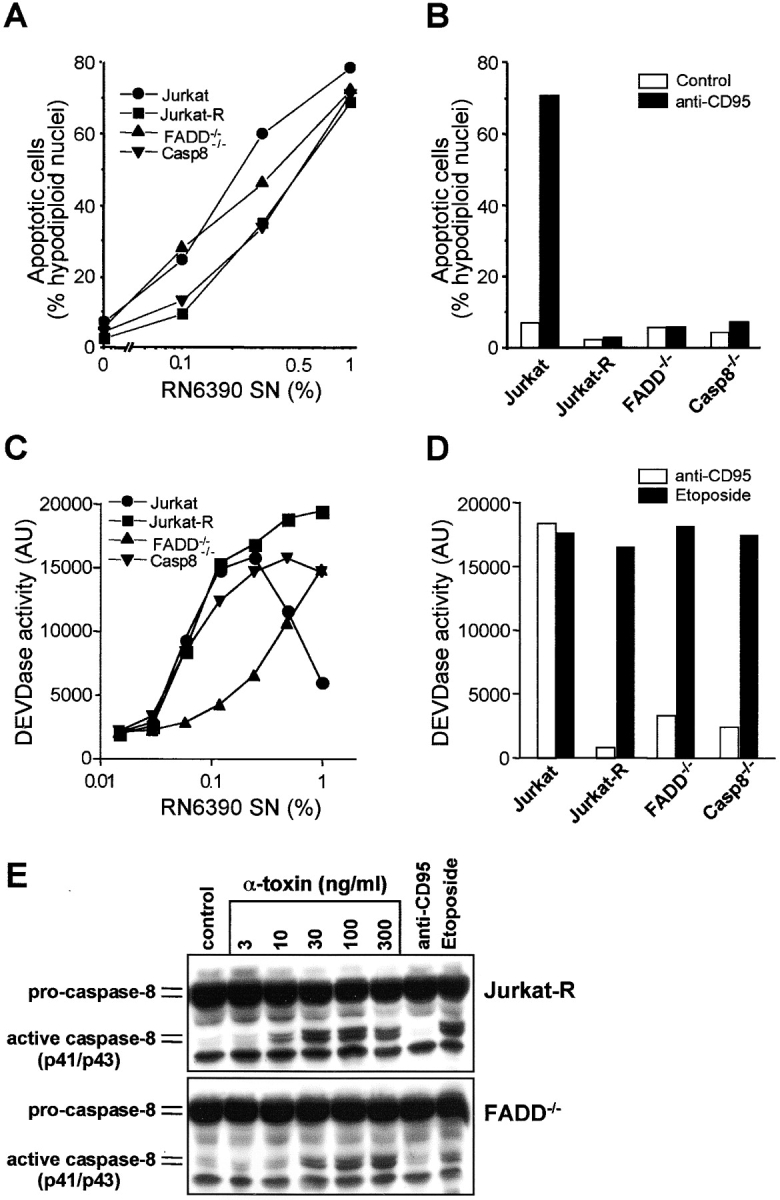
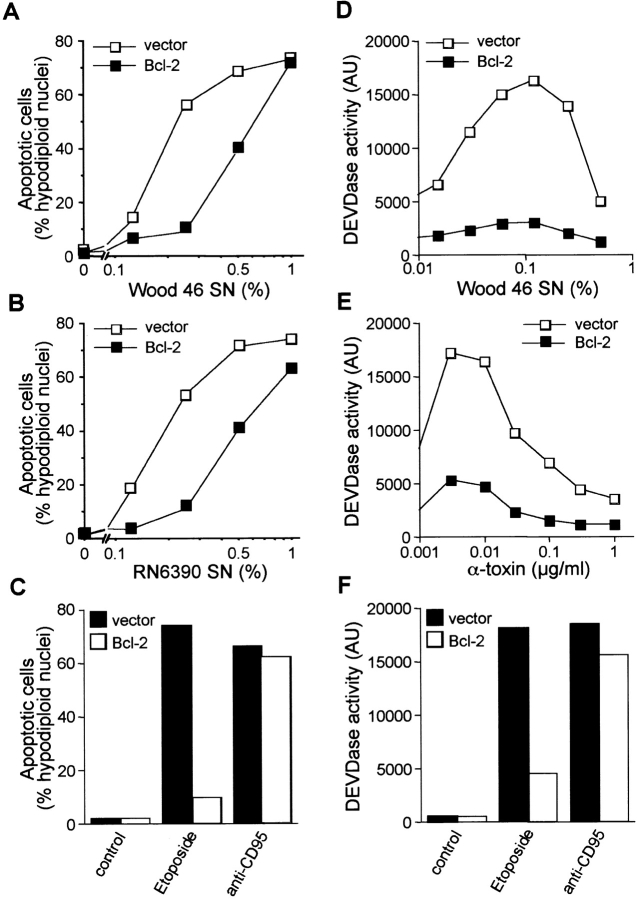
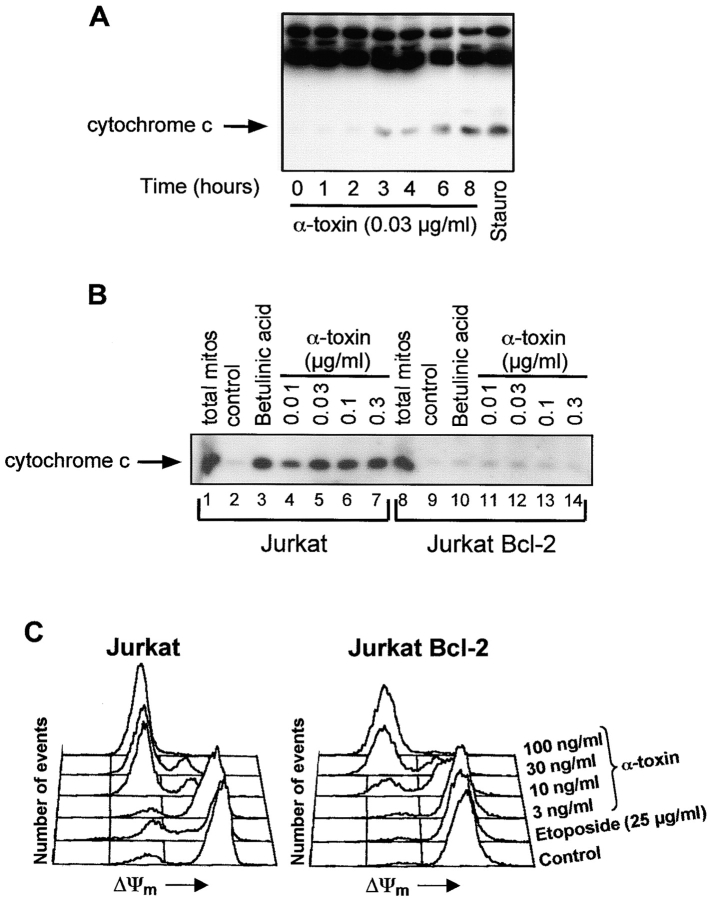
Infections with Staphylococcus aureus, a common inducer of septic and toxic shock, often result in tissue damage and death of various cell types. Although S. aureus was suggested to induce apoptosis, the underlying signal transduction pathways remained elusive. We show that caspase activation and DNA fragmentation were induced not only when Jurkat T cells were infected with intact bacteria, but also after treatment with supernatants of various S. aureus strains. We also demonstrate that S. aureus-induced cell death and caspase activation were mediated by alpha-toxin, a major cytotoxin of S. aureus, since both events were abrogated by two different anti-alpha-toxin antibodies and could not be induced with supernatants of an alpha-toxin-deficient S. aureus strain. Furthermore, alpha-toxin-induced caspase activation in CD95-resistant Jurkat sublines lacking CD95, Fas-activated death domain, or caspase-8 but not in cells stably expressing the antiapoptotic protein Bcl-2. Together with our finding that alpha-toxin induces cytochrome c release in intact cells and, interestingly, also from isolated mitochondria in a Bcl-2-controlled manner, our results demonstrate that S. aureus alpha-toxin triggers caspase activation via the intrinsic death pathway independently of death receptors. Hence, our findings clearly define a signaling pathway used in S. aureus-induced cytotoxicity and may provide a molecular rationale for future therapeutic interventions in bacterial infections.
Figures
References
-
- Balwit, J.M., P. van Langevelde, J.M. Vann, and R.A. Proctor. 1994. Gentamicin-resistant menadione and hemin auxotrophic Staphylococcus aureus persist within cultured endothelial cells. J. Infect. Dis. 170:1033–1037. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
