

Antibacterial effect of human V gamma 2V delta 2 T cells in vivo
- PMID: 11696580
- PMCID: PMC209444
- DOI: 10.1172/JCI13584
Antibacterial effect of human V gamma 2V delta 2 T cells in vivo
Abstract
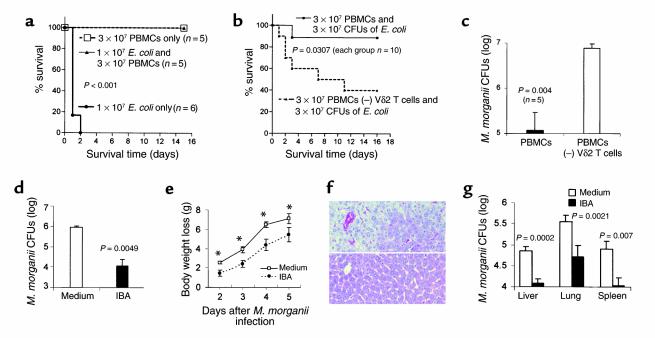
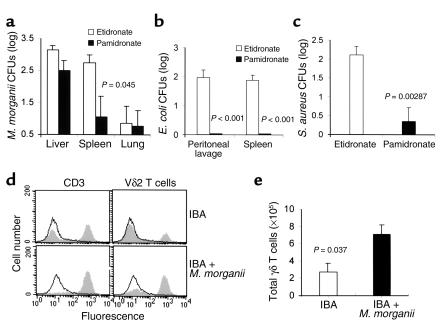
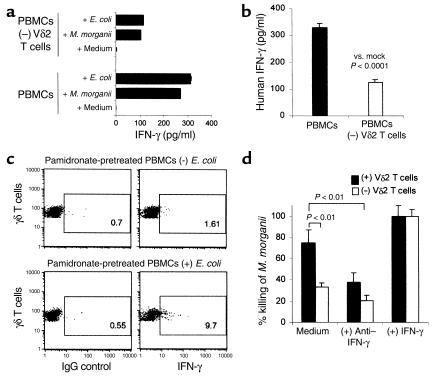
V gamma 2V delta 2 cells, a class of T cells found only in primates, are reactive to nonpeptide organophosphate and alkylamine antigens secreted by bacteria and parasites. These cells make up 2-5% percent of human peripheral blood T cells but expand to make up 8-60% of peripheral blood T cells during bacterial and parasitic infections. We show here, using a chimeric severe combined immunodeficiency (SCID) mouse (hu-SCID) model, that human V gamma 2V delta 2 T cells mediate resistance to extracellular gram-positive (Staphylococcus aureus) and gram-negative (Escherichia coli and Morganella morganii) bacteria, as assessed by survival, body weight, bacterial loads, and histopathology. Surprisingly, this bacterial resistance was evident 1 day after infection, and bacteria were cleared well before gamma delta T cell expansion was detected 6 days after infection. Decreased resistance in V delta 2 T cell-depleted hu-SCID mice correlated with decreased serum IFN-gamma titers. Intravenous treatment of infected, reconstituted hu-SCID mice with pamidronate, a human V gamma 2V delta 2 T cell-specific aminobisphosphonate antigen, markedly increased the in vivo antibacterial effect of V gamma 2V delta 2 T cells. Therefore, this large pool of antigen-specific, yet immediately reactive memory human V gamma 2V delta 2 T cells is likely to be an important mediator of resistance against extracellular bacterial infection and may bridge the gap between innate and acquired immunity.
Figures
References
-
- Ladel CH, Blum C, Dreher A, Reifenberg K, Kaufmann SH. Protective role of γδ T cells and αβ T cells in tuberculosis. Eur J Immunol. 1995;25:2877–2881. - PubMed
-
- Peterman GM, Spencer C, Sperling AI, Bluestone JA. Role of γδ T cells in murine collagen-induced arthritis. J Immunol. 1993;151:6546–6558. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
