

Gap junctional coupling in lenses from alpha(8) connexin knockout mice
- PMID: 11696604
- PMCID: PMC2233836
- DOI: 10.1085/jgp.118.5.447
Gap junctional coupling in lenses from alpha(8) connexin knockout mice
Abstract
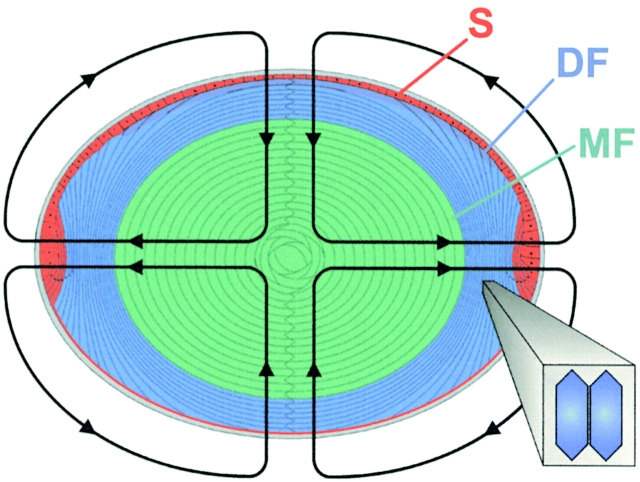
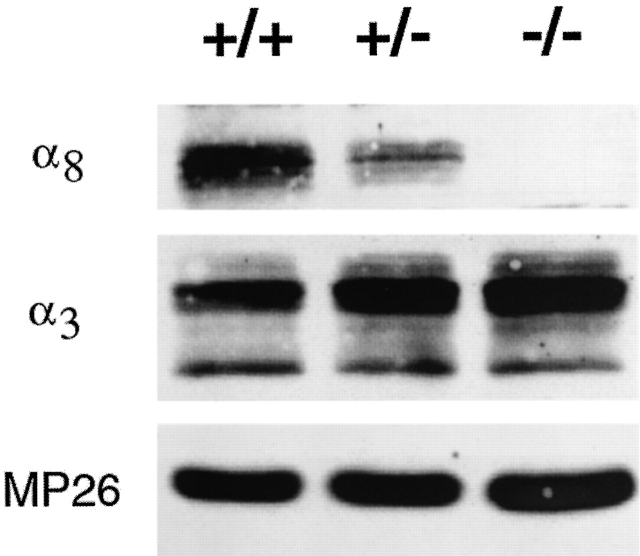
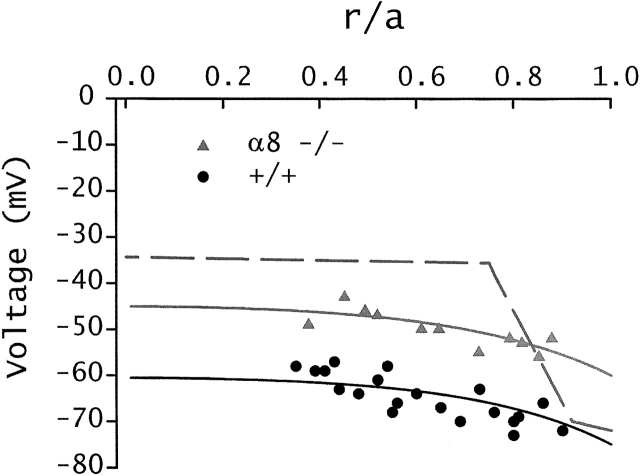
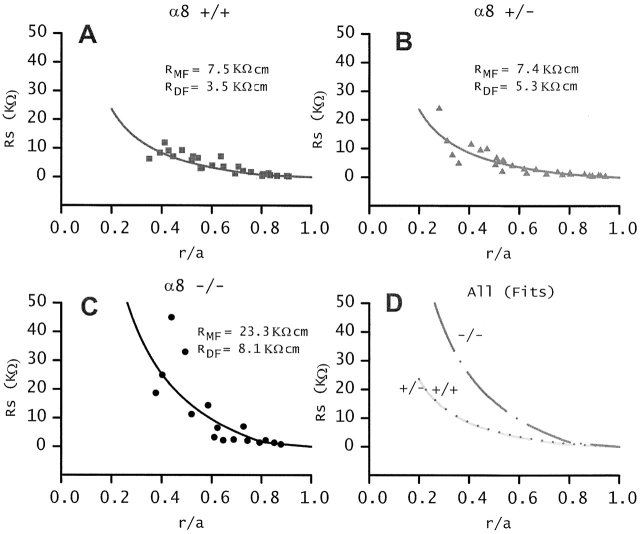
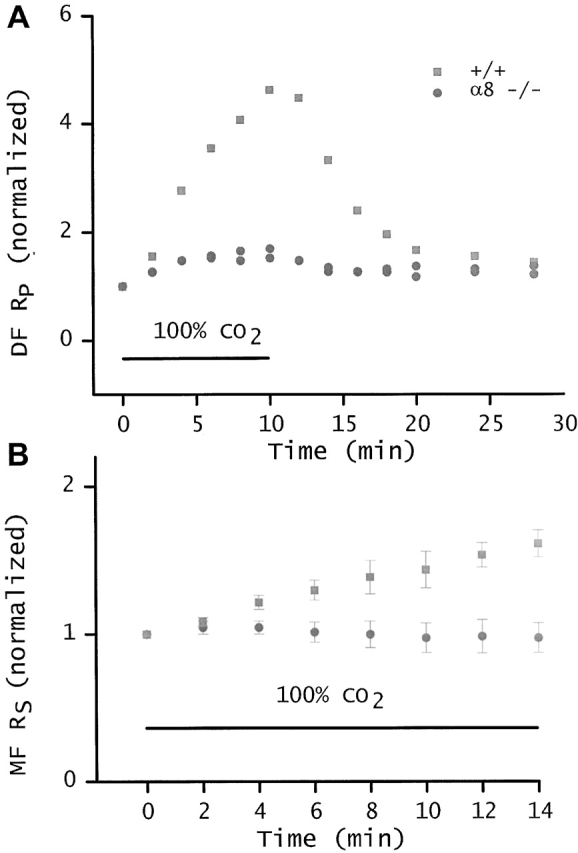
Lens fiber cell gap junctions contain alpha(3) (Cx46) and alpha(8) (Cx50) connexins. To examine the roles of the two different connexins in lens physiology, we have genetically engineered mice lacking either alpha(3) or alpha(8) connexin. Intracellular impedance studies of these lenses were used to measure junctional conductance and its sensitivity to intracellular pH. In Gong et al. 1998, we described results from alpha(3) connexin knockout lenses. Here, we present original data from alpha(8) connexin knockout lenses and a comparison with the previous results. The lens has two functionally distinct domains of fiber cell coupling. In wild-type mouse lenses, the outer shell of differentiating fibers (see 1, DF) has an average coupling conductance per area of cell-cell contact of approximately 1 S/cm(2), which falls to near zero when the cytoplasm is acidified. In the inner core of mature fibers (see 1, MF), the average coupling conductance is approximately 0.4 S/cm(2), and is insensitive to acidification of the cytoplasm. Both connexin isoforms appear to contribute about equally in the DF since the coupling conductance for either heterozygous knockout (+/-) was approximately 70% of normal and 30-40% of the normal for both -/- lenses. However, their contribution to the MF was different. About 50% of the normal coupling conductance was found in the MF of alpha(3) +/- lenses. In contrast, the coupling of MF in the alpha(8) +/- lenses was the same as normal. Moreover, no coupling was detected in the MF of alpha(3) -/- lenses. Together, these results suggest that alpha(3) connexin alone is responsible for coupling MF. The pH- sensitive gating of DF junctions was about the same in wild-type and alpha(3) connexin -/- lenses. However, in alpha(8) -/- lenses, the pure alpha(3) connexin junctions did not gate closed in the response to acidification. Since alpha(3) connexin contributes about half the coupling conductance in DF of wild-type lenses, and that conductance goes to zero when the cytoplasmic pH drops, it appears alpha(8) connexin regulates the gating of alpha(3) connexin. Both connexins are clearly important to lens physiology as lenses null for either connexin lose transparency. Gap junctions in the MF survive for the lifetime of the organism without protein turnover. It appears that alpha(3) connexin provides the long-term communication in MF. Gap junctions in DF may be physiologically regulated since they are capable of gating when the cytoplasm is acidified. It appears alpha(8) connexin is required for gating in DF.
Figures
References
-
- Bassnett S., Croghan P.C., Duncan G. Diffusion of lactate and its role in determining intracellular pH in the lens of the eye. Exp. Eye Res. 1987;44:143–147. - PubMed
-
- Donaldson P., Kistler J., Mathias R.T. Molecular solutions to lens transparency. News Physiol. Sci. 2001;16:118–123. - PubMed
-
- Fischbarg J., Diecke F.P., Kuang K., Yu B., Kang F., Iserovich P., Li Y., Rosskothen H., Koniarek J.P. Transport of fluid by lens epithelium. Am. J. Physiol. 1999;276:C548–C557. - PubMed
-
- Gao J., Sun X., Yatsula V., Wymore R.S., Mathias R.T. Isoform-specific function and distribution of Na/K pumps in the frog lens epithelium. J. Membr. Biol. 2000;178:89–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
