

Anatomical distribution and cellular basis for high levels of aromatase activity in the brain of teleost fish: aromatase enzyme and mRNA expression identify glia as source
- PMID: 11698605
- PMCID: PMC6762278
- DOI: 10.1523/JNEUROSCI.21-22-08943.2001
Anatomical distribution and cellular basis for high levels of aromatase activity in the brain of teleost fish: aromatase enzyme and mRNA expression identify glia as source
Abstract
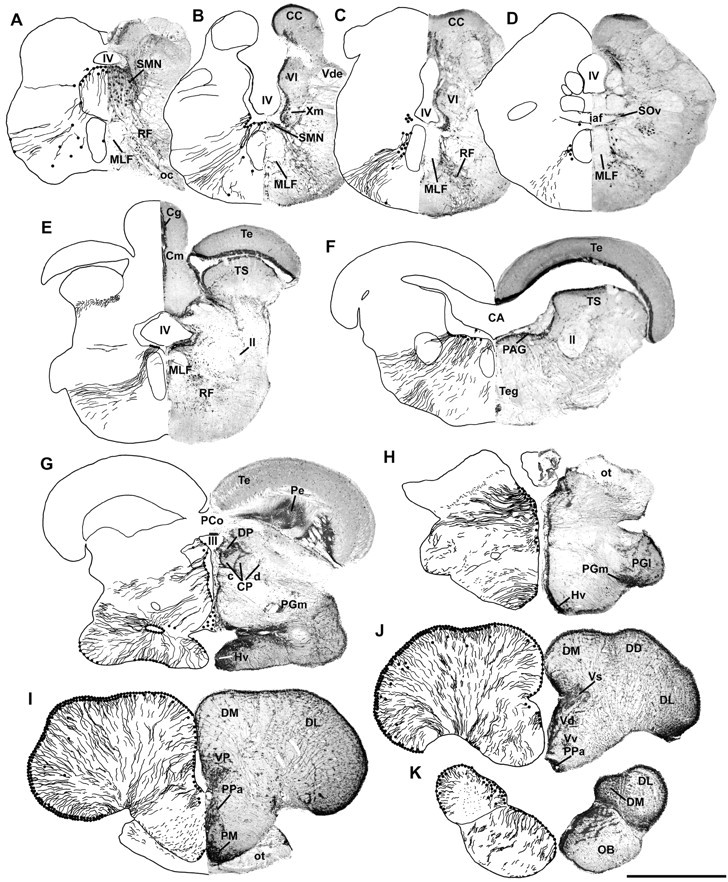
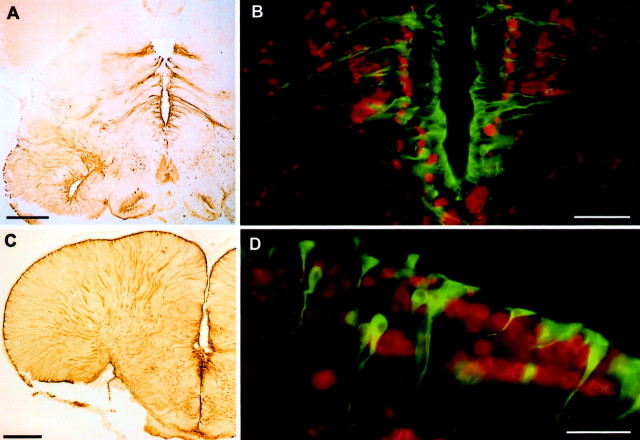
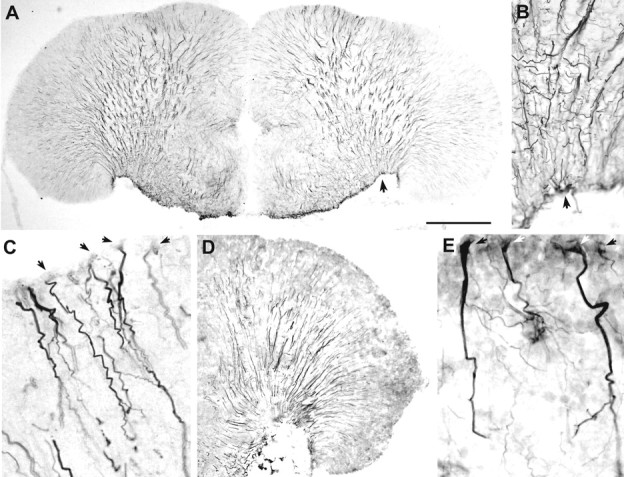
Although teleost fish have higher levels of brain aromatase activity than any other vertebrate group, its function remains speculative, and no study has identified its cellular basis. A previous study determined aromatase activity in a vocal fish, the plainfin midshipman (Porichthys notatus), and found highest levels in the telencephalon and lower levels in the sonic hindbrain, which was dimorphic between and within (males) sexes. We have now localized aromatase-containing cells in the midshipman brain both by immunocytochemistry using teleost-specific aromatase antibodies and by in situ hybridization using midshipman-specific aromatase probes. Aromatase-immuno-reactivity and mRNA hybridization signal are consistent with relative levels of aromatase activity in different brain regions: concentrated in the dimorphic sonic motor nucleus, in a band just beneath the periaqueductal gray in the midbrain, in ventricular regions in the hypothalamus, and highest levels in the telencephalon especially in preoptic and ventricular areas. Surprisingly, double-label immunofluorescence does not show aromatase-immunoreactive colocalization in neurons, but instead in radial glia throughout the brain. This is the first study to identify aromatase expression mostly, if not entirely, in glial cells under normal rather than brain injury-dependent conditions. The abundance of aromatase in teleosts may represent an adaptation linked to continual neurogenesis that is known to occur throughout an individual's lifetime among fishes. The localization of aromatase within the intersexually and intrasexually dimorphic vocal-motor circuit further implies a function in the expression of alternative male reproductive phenotypes and, more generally, the development of natural, individual variation of specific brain nuclei.
Figures






Similar articles
-
Seasonal plasticity of brain aromatase mRNA expression in glia: divergence across sex and vocal phenotypes.J Neurobiol. 2005 Oct;65(1):37-49. doi: 10.1002/neu.20179. J Neurobiol. 2005. PMID: 16003720
-
Distribution of androgen receptor mRNA expression in vocal, auditory, and neuroendocrine circuits in a teleost fish.J Comp Neurol. 2010 Feb 15;518(4):493-512. doi: 10.1002/cne.22233. J Comp Neurol. 2010. PMID: 20020540 Free PMC article.
-
Sexually-dimorphic expression of tyrosine hydroxylase immunoreactivity in the brain of a vocal teleost fish (Porichthys notatus).J Chem Neuroanat. 2014 Mar;56:13-34. doi: 10.1016/j.jchemneu.2014.01.001. Epub 2014 Jan 10. J Chem Neuroanat. 2014. PMID: 24418093
-
Brain aromatase: new lessons from non-mammalian model systems.Front Neuroendocrinol. 2006 Sep;27(3):247-74. doi: 10.1016/j.yfrne.2006.05.002. Epub 2006 Jul 10. Front Neuroendocrinol. 2006. PMID: 16828853 Review.
-
Relationships between aromatase and estrogen receptors in the brain of teleost fish.Gen Comp Endocrinol. 2005 May 15;142(1-2):60-6. doi: 10.1016/j.ygcen.2004.12.003. Epub 2005 Jan 20. Gen Comp Endocrinol. 2005. PMID: 15862549 Review.
Cited by
-
Brain estrogen signaling effects acute modulation of acoustic communication behaviors: A working hypothesis.Bioessays. 2012 Dec;34(12):1009-16. doi: 10.1002/bies.201200081. Epub 2012 Oct 15. Bioessays. 2012. PMID: 23065844 Free PMC article.
-
Localization and divergent profiles of estrogen receptors and aromatase in the vocal and auditory networks of a fish with alternative mating tactics.J Comp Neurol. 2013 Aug 15;521(12):2850-69. doi: 10.1002/cne.23320. J Comp Neurol. 2013. PMID: 23460422 Free PMC article.
-
Aromatase immunoreactivity in the bluehead wrasse brain, Thalassoma bifasciatum: immunolocalization and co-regionalization with arginine vasotocin and tyrosine hydroxylase.Brain Res. 2006 Dec 18;1126(1):91-101. doi: 10.1016/j.brainres.2006.09.017. Epub 2006 Oct 11. Brain Res. 2006. PMID: 17045250 Free PMC article.
-
Widespread capacity for steroid synthesis in the avian brain and song system.Endocrinology. 2006 Dec;147(12):5975-87. doi: 10.1210/en.2006-0154. Epub 2006 Aug 24. Endocrinology. 2006. PMID: 16935847 Free PMC article.
-
Local expression of CYP19A1 and CYP19A2 in developing and adult killifish (Fundulus heteroclitus).Gen Comp Endocrinol. 2008 Jan 15;155(2):307-17. doi: 10.1016/j.ygcen.2007.05.018. Epub 2007 May 21. Gen Comp Endocrinol. 2008. PMID: 17582409 Free PMC article.
References
-
- Alvarez-Buylla A, Kirn J. Birth, migration, incorporation, and death of vocal control neurons in adult songbirds. J Neurobiol. 1997;33:585–601. - PubMed
-
- Alvarez-Buylla A, Buskirk DR, Nottebohm F. Monoclonal antibody reveals radial glia in adult avian brain. J Comp Neurol. 1987;264:159–170. - PubMed
-
- Alvarez-Buylla A, Garcia-Verdugo JM, Tramontin AD. A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci. 2001;2:287–293. - PubMed
-
- Balthazart J, Ball G. New insights into the regulation and function of brain estrogen synthase (aromatase). Trends Neurosci. 1998;21:243–248. - PubMed
-
- Bass AH. Dimorphic male brains and alternate reproductive tactics in a vocalizing fish. Trends Neurosci. 1992;15:139–145. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources