

Combined effects of intrinsic facilitation and modulatory inhibition of identified interneurons in the siphon withdrawal circuitry of Aplysia
- PMID: 11698609
- PMCID: PMC6762296
- DOI: 10.1523/JNEUROSCI.21-22-08990.2001
Combined effects of intrinsic facilitation and modulatory inhibition of identified interneurons in the siphon withdrawal circuitry of Aplysia
Abstract
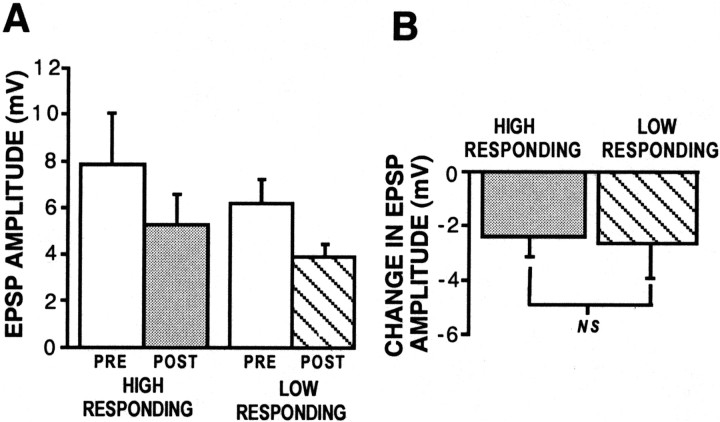
Synaptic plasticity can be induced through mechanisms intrinsic to a synapse or through extrinsic modulatory mechanisms. In this study, we investigated the relationship between these two forms of plasticity at the excitatory synapse between L29 interneurons and siphon motor neurons (MNs) in Aplysia. Using isolated ganglia, we confirmed that the L29-MN synapses exhibit a form of intrinsic facilitation: post-tetanic potentiation (PTP). We also found that L29-MN synapses are modulated by exogenous application of 5-HT: they are depressed after 5-HT exposure. We next investigated the functional relationship between an intrinsic facilitatory process (PTP) and extrinsic inhibitory modulation (5-HT-induced depression). First, we found that application of 5-HT just before L29 activation results in a reduction of PTP. Second, using semi-intact preparations, we found that tail shock (TS) mimics the effect of 5-HT by both depressing L29 synaptic transmission and by reducing L29 PTP. Third, we observed a significant correlation between L29 activity during TS and subsequent synaptic change: low-responding L29s showed synaptic depression after TS, whereas high-responding L29s showed synaptic facilitation. Finally, we found that we could directly manipulate the sign and magnitude of TS-induced synaptic plasticity by controlling L29 activity during TS. Collectively, our results show that the L29-MN synapses exhibit intrinsic facilitation and extrinsic modulation and that the sign and magnitude of L29-MN plasticity induced by TS is governed by the combined effects of these two processes. This circuit architecture, which combines network inhibition with cell-specific facilitation, can enhance the signal value of a specific stimulus within a neural network.
Figures










Similar articles
-
Contribution of postsynaptic Ca2+ to the induction of post-tetanic potentiation in the neural circuit for siphon withdrawal in Aplysia.J Neurosci. 2001 Mar 1;21(5):1739-49. doi: 10.1523/JNEUROSCI.21-05-01739.2001. J Neurosci. 2001. PMID: 11222663 Free PMC article.
-
Sensitization of the gill and siphon withdrawal reflex of Aplysia: multiple sites of change in the neuronal network.J Neurophysiol. 1993 Sep;70(3):1210-20. doi: 10.1152/jn.1993.70.3.1210. J Neurophysiol. 1993. PMID: 8229169
-
Modeling of substance P and 5-HT induced synaptic plasticity in the lamprey spinal CPG: consequences for network pattern generation.J Comput Neurosci. 2001 Sep-Oct;11(2):183-200. doi: 10.1023/a:1012806018730. J Comput Neurosci. 2001. PMID: 11717534
-
Functional roles of short-term synaptic plasticity with an emphasis on inhibition.Curr Opin Neurobiol. 2017 Apr;43:71-78. doi: 10.1016/j.conb.2017.01.002. Epub 2017 Jan 22. Curr Opin Neurobiol. 2017. PMID: 28122326 Free PMC article. Review.
-
Synapse regression in depression: the role of 5-HT receptors in modulating NMDA receptor function and synaptic plasticity.Aust N Z J Psychiatry. 2010 Apr;44(4):301-8. doi: 10.3109/00048670903555146. Aust N Z J Psychiatry. 2010. PMID: 20307163 Review.
Cited by
-
Parallel somatic and synaptic processing in the induction of intermediate-term and long-term synaptic facilitation in Aplysia.Proc Natl Acad Sci U S A. 2004 May 11;101(19):7463-8. doi: 10.1073/pnas.0402163101. Epub 2004 Apr 29. Proc Natl Acad Sci U S A. 2004. PMID: 15123836 Free PMC article.
-
Regulation of behavioral and synaptic plasticity by serotonin release within local modulatory fields in the CNS of Aplysia.J Neurosci. 2006 Dec 6;26(49):12682-93. doi: 10.1523/JNEUROSCI.3309-06.2006. J Neurosci. 2006. PMID: 17151271 Free PMC article.
-
Multiple serotonergic mechanisms contributing to sensitization in aplysia: evidence of diverse serotonin receptor subtypes.Learn Mem. 2003 Sep-Oct;10(5):373-86. doi: 10.1101/lm.66103. Learn Mem. 2003. PMID: 14557610 Free PMC article. Review.
-
Evolution of learning in three aplysiid species: differences in heterosynaptic plasticity contrast with conservation in serotonergic pathways.J Physiol. 2003 Jul 1;550(Pt 1):241-53. doi: 10.1113/jphysiol.2003.038356. Epub 2003 May 9. J Physiol. 2003. PMID: 12740422 Free PMC article.
-
Peptide neuromodulation of synaptic dynamics in an oscillatory network.J Neurosci. 2011 Sep 28;31(39):13991-4004. doi: 10.1523/JNEUROSCI.3624-11.2011. J Neurosci. 2011. PMID: 21957260 Free PMC article.
References
-
- Barbeau H, Rossignol S. Initiation and modulation of the locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic, and dopaminergic drugs. Brain Res. 1991;546:250–260. - PubMed
-
- Belkin KJ, Abrams TW. The effect of the neuropeptide FMRFamide on Aplysia californica siphon motoneurons involves multiple ionic currents that vary seasonally. J Exp Biol. 1998;201:2225–2234. - PubMed
-
- Bristol AS, Fischer TM, Carew TJ. Characterization and modulation of activity-dependent plasticity at the L29-MN synapse in Aplysia. Soc Neurosci Abstr. 1998;24:702.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources