

Cloning, expression, and regulation of a glucocorticoid-induced receptor in rat brain: effect of repetitive amphetamine
- PMID: 11698613
- PMCID: PMC6762284
- DOI: 10.1523/JNEUROSCI.21-22-09027.2001
Cloning, expression, and regulation of a glucocorticoid-induced receptor in rat brain: effect of repetitive amphetamine
Abstract
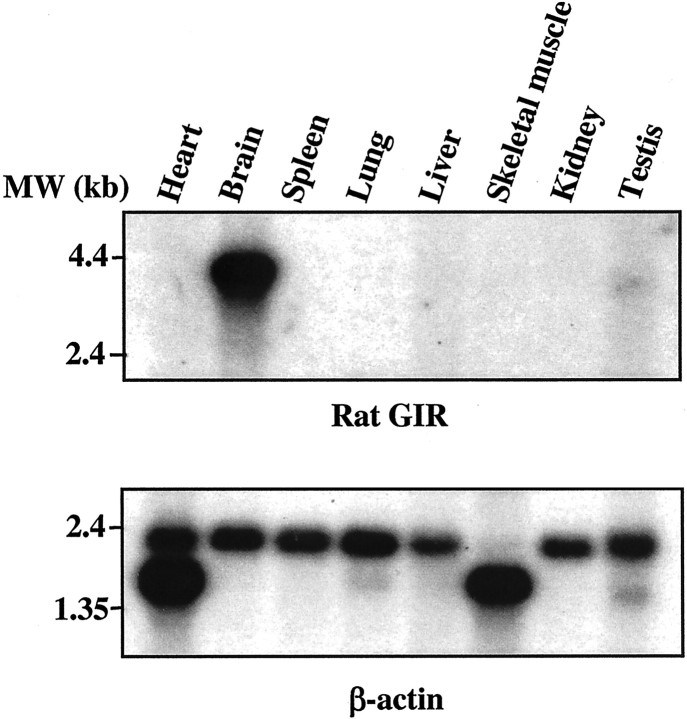
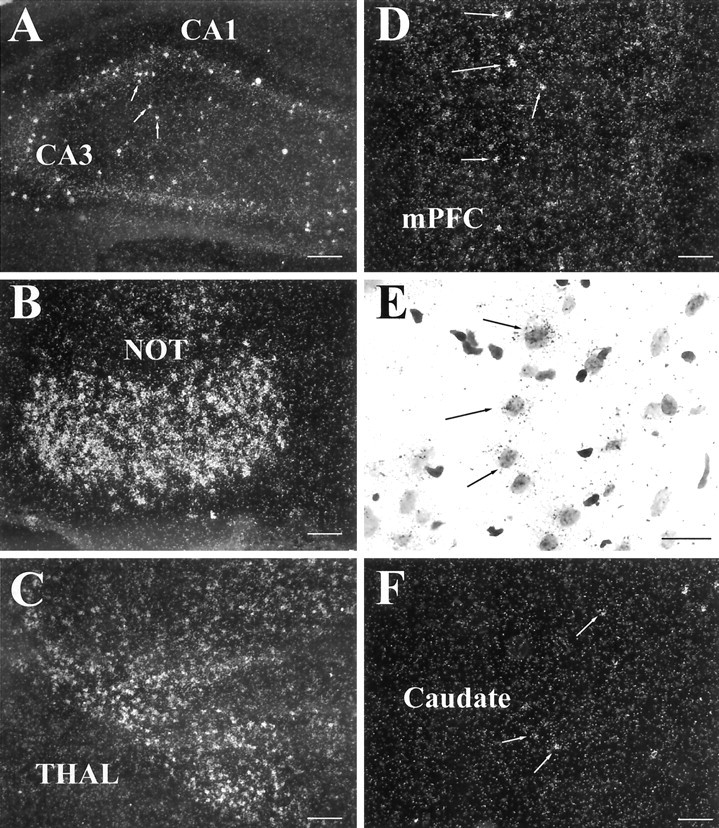
Behavioral sensitization to psychostimulants involves neuroadaptation of stress-responsive systems. We have identified and sequenced a glucocorticoid-induced receptor (GIR) cDNA from rat prefrontal cortex. The full-length GIR cDNA encodes a 422 amino acid protein belonging to G-protein-coupled receptor superfamily. Although the ligand for GIR is still unknown, the dendrogram construction indicates that GIR may belong to peptide receptor subfamily (e.g., substance P receptor), with more distant relationship to subfamilies of glycoprotein hormone receptors (e.g., thyrotropin receptor) and biogenic amine receptors (e.g., dopamine receptor). GIR shares 31-34% amino acid identity to the tachykinin receptors (substance P receptor, neurokinin A receptor, and neurokinin B receptor). GIR mRNA is expressed preferentially in brain, and its neuronal expression is relegated to limbic brain regions, particularly in forebrain. GIR transcript levels are increased significantly and persistently in prefrontal cortex for 7 d after discontinuation of chronic amphetamine exposure. The induction of GIR expression by amphetamine is associated with augmented behavioral activation. These findings suggest that modulation of GIR expression may be involved in behavioral sensitization, and GIR may play a role at the interface between stress and neuroadaptation to psychostimulants.
Figures




References
-
- Antelman SM, Eichler AJ, Black CA, Kocan D. Interchangeability of stress and amphetamine in sensitization. Science. 1980;207:329–331. - PubMed
-
- Bouvier M, Collins S, O'Dowd BF, Campbell PT, de Blasi A, Kobilka BK, MacGregor C, Irons GP, Caron MG, Lefkowitz RJ. Two distinct pathways for cAMP-mediated down-regulation of the beta 2-adrenergic receptor. Phosphorylation of the receptor and regulation of its mRNA level. J Biol Chem. 1989;264:16786–16792. - PubMed
-
- Casada JH, Dafny N. Restraint and stimulation of the bed nucleus of the stria terminalis produce similar stress-like behaviors. Brain Res Bull. 1991;27:207–212. - PubMed
-
- Clark RB, Friedman J, Dixon RA, Strader CD. Identification of a specific site required for rapid heterologous desensitization of the beta-adrenergic receptor by cAMP-dependent protein kinase. Mol Pharmacol. 1989;36:343–348. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases