

Regulation of CAX1, an Arabidopsis Ca(2+)/H+ antiporter. Identification of an N-terminal autoinhibitory domain
- PMID: 11706183
- PMCID: PMC129272
Regulation of CAX1, an Arabidopsis Ca(2+)/H+ antiporter. Identification of an N-terminal autoinhibitory domain
Abstract
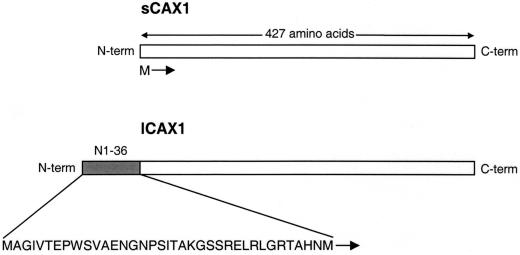
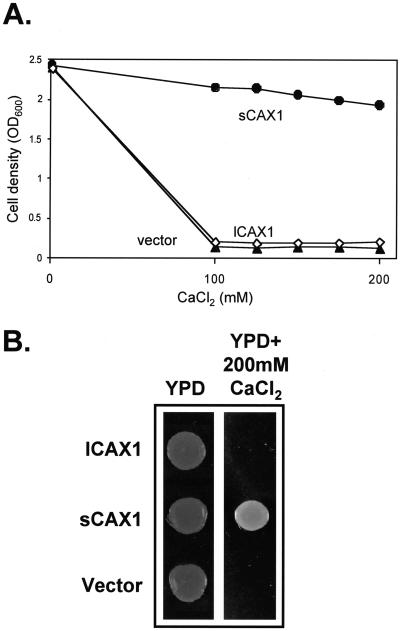
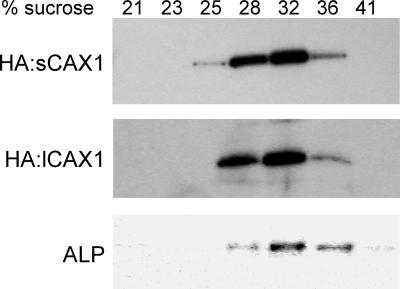
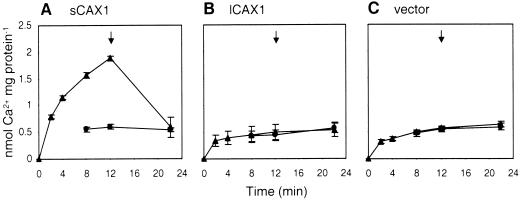
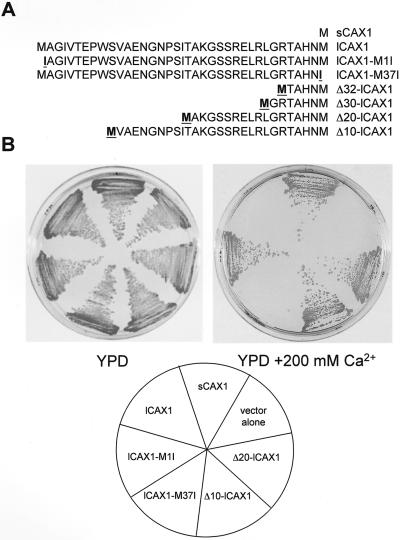
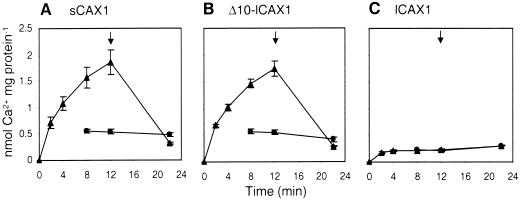
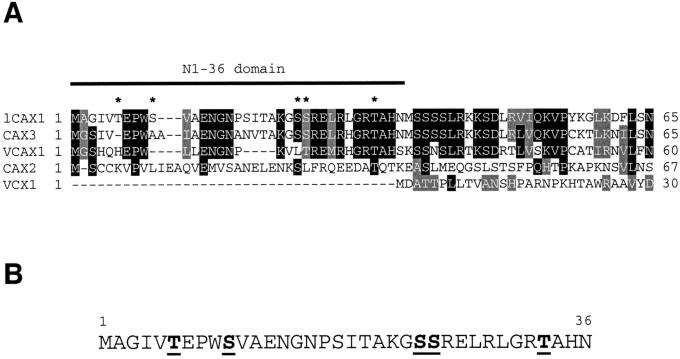
Regulation of Ca(2+) transport determines the duration of a Ca(2+) signal, and hence, the nature of the biological response. Ca(2+)/H+ antiporters such as CAX1 (cation exchanger 1), play a key role in determining cytosolic Ca(2+) levels. Analysis of a full-length CAX1 clone suggested that the CAX1 open reading frame contains an additional 36 amino acids at the N terminus that were not found in the original clone identified by suppression of yeast (Saccharomyces cerevisiae) vacuolar Ca(2+) transport mutants. The long CAX1 (lCAX1) could not suppress the yeast Ca(2+) transport defects despite localization to the yeast vacuole. Calmodulin could not stimulate lCAX1 Ca(2+)/H+ transport in yeast; however, minor alterations in the 36-amino acid region restored Ca(2+)/H+ transport. Sequence analysis suggests that a 36-amino acid N-terminal regulatory domain may be present in all Arabidopsis CAX-like genes. Together, these results suggest a structural feature involved in regulation of Ca(2+)/H+ antiport.
Figures
References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. New York: Greene Publishing Associates/Wiley Interscience; 1998.
-
- Blom N, Gammeltoft S, Brunak S. Sequence- and structure-based prediction of eukaryotic protein phosphorylation sites. J Mol Biol. 1999;294:1351–1362. - PubMed
-
- Bush DS. Calcium regulation in plant cells and its role in signaling. Annu Rev Plant Physiol Plant Mol Biol. 1995;46:95–122.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous