

A gene controlling variation in Arabidopsis glucosinolate composition is part of the methionine chain elongation pathway
- PMID: 11706188
- PMCID: PMC129277
A gene controlling variation in Arabidopsis glucosinolate composition is part of the methionine chain elongation pathway
Abstract
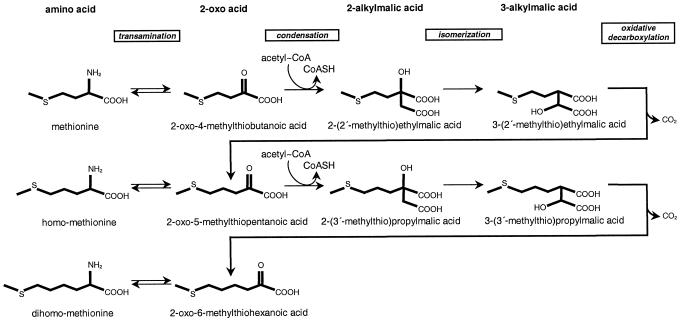
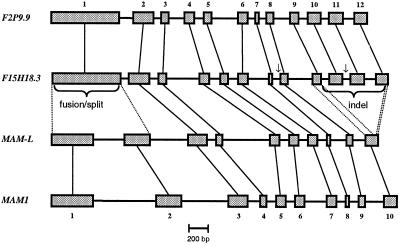
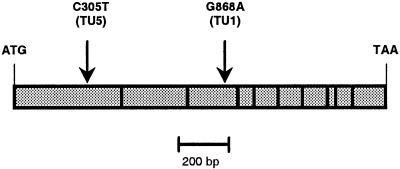
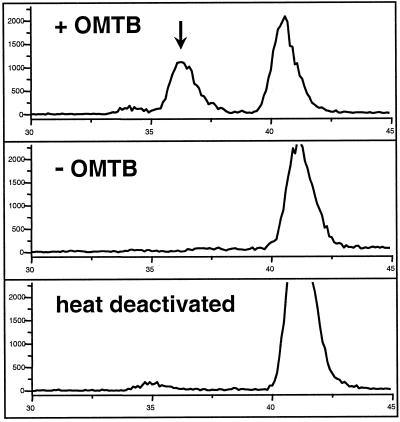
Arabidopsis and other Brassicaceae produce an enormous diversity of aliphatic glucosinolates, a group of methionine (Met)-derived plant secondary compounds containing a beta-thio-glucose moiety, a sulfonated oxime, and a variable side chain. We fine-scale mapped GSL-ELONG, a locus controlling variation in the side-chain length of aliphatic glucosinolates. Within this locus, a polymorphic gene was identified that determines whether Met is extended predominantly by either one or by two methylene groups to produce aliphatic glucosinolates with either three- or four-carbon side chains. Two allelic mutants deficient in four-carbon side-chain glucosinolates were shown to contain independent missense mutations within this gene. In cell-free enzyme assays, a heterologously expressed cDNA from this locus was capable of condensing 2-oxo-4-methylthiobutanoic acid with acetyl-coenzyme A, the initial reaction in Met chain elongation. The gene methylthioalkylmalate synthase1 (MAM1) is a member of a gene family sharing approximately 60% amino acid sequence similarity with 2-isopropylmalate synthase, an enzyme of leucine biosynthesis that condenses 2-oxo-3-methylbutanoate with acetyl-coenzyme A.
Figures


References
-
- Blau PA, Feeny P, Contardo L. Allylglucosinolate and herbivorous caterpillars: a contrast in toxicity and tolerance. Science. 1978;200:1296–1298. - PubMed
-
- Bodnaryk RP. Developmental profile of sinalbin (p-hydroxybenzyl glucosinolate) in mustard seedlings, Sinapis albaL., and its relationship to insect resistance. J Chem Ecol. 1991;17:1543–1556. - PubMed
-
- Bones AM, Rossiter JT. The myrosinase-glucosinolate system, its organization and biochemistry. Physiol Plant. 1996;97:194–208.
-
- Bradshaw JE, Heaney RK, Smith WHM, Gowers S, Gemmell DJ, Fenwick GR. The glucosinolate content of some fodder brassicas. J Sci Food Agric. 1984;35:977–981.
-
- Brown PD, Morra MJ. Glucosinolate-containing plant tissues as bioherbicides. J Agric Food Chem. 1995;43:3070–3074.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous