

Hyperactivation of the yeast DNA damage checkpoint by TEL1 and DDC2 overexpression
- PMID: 11707419
- PMCID: PMC125310
- DOI: 10.1093/emboj/20.22.6485
Hyperactivation of the yeast DNA damage checkpoint by TEL1 and DDC2 overexpression
Abstract
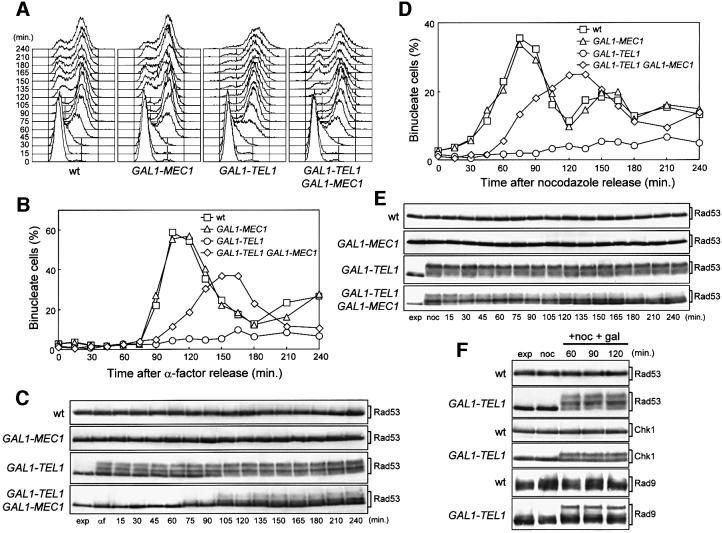
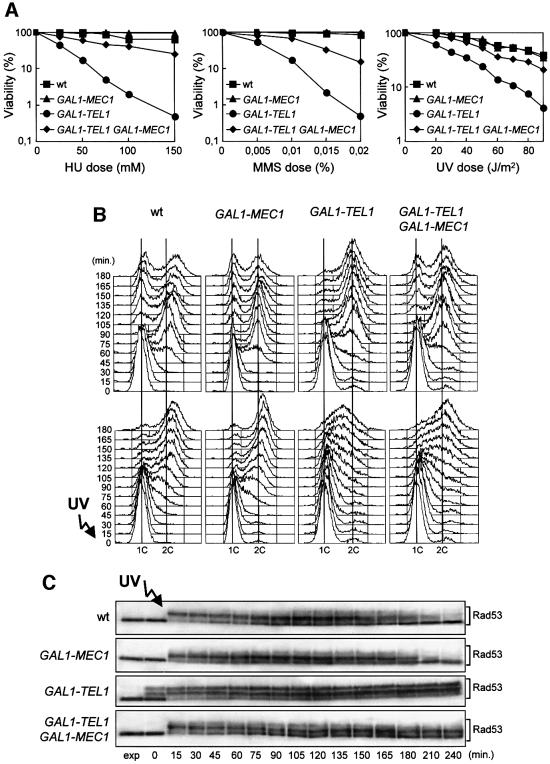
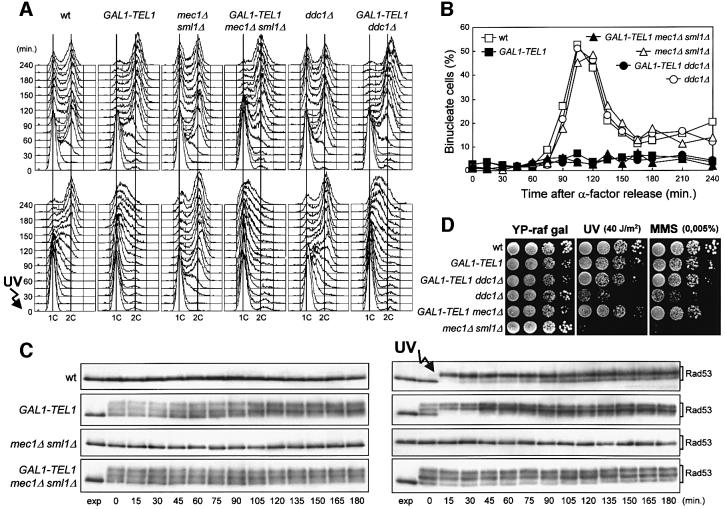
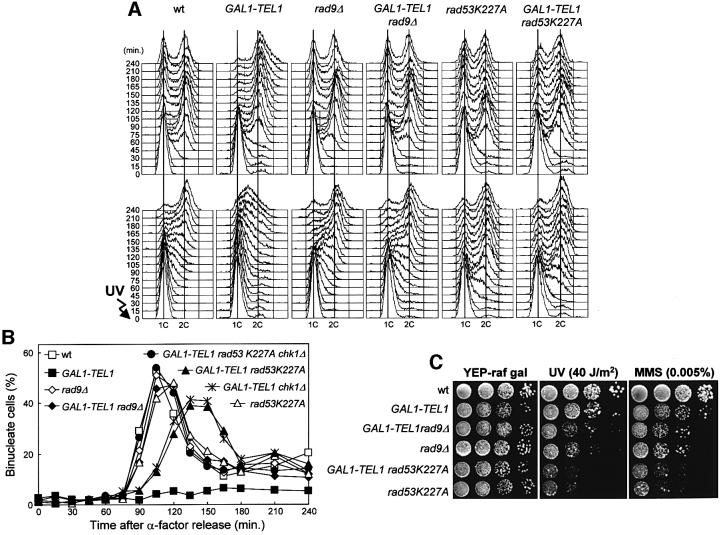
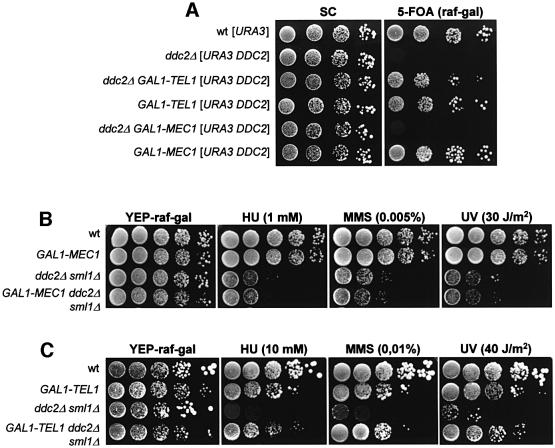
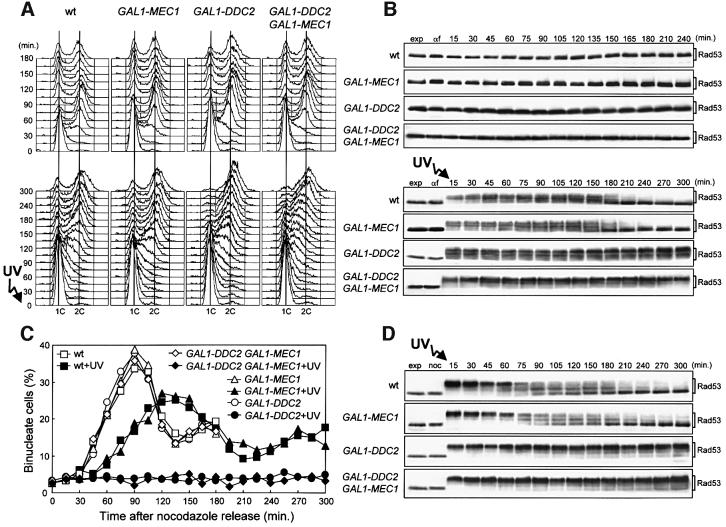
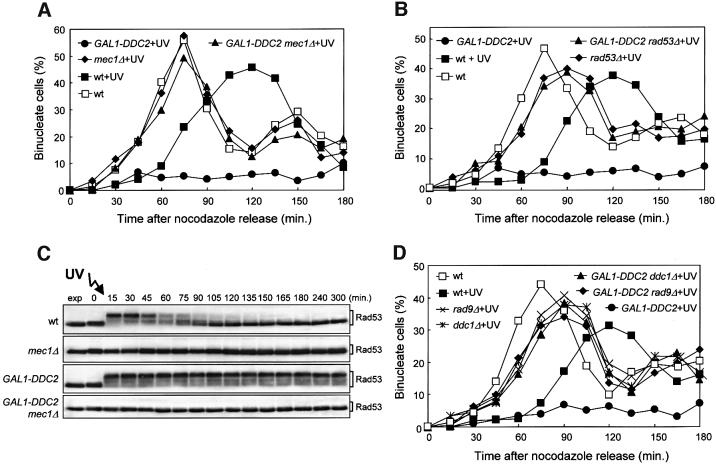
The evolutionarily conserved yeast Mec1 and Tel1 protein kinases, as well as the Mec1-interacting protein Ddc2, are involved in the DNA damage checkpoint response. We show that regulation of Tel1 and Ddc2-Mec1 activities is important to modulate both activation and termination of checkpoint-mediated cell cycle arrest. In fact, overproduction of either Tel1 or Ddc2 causes a prolonged cell cycle arrest and cell death in response to DNA damage, impairing the ability of cells to recover from checkpoint activation. This cell cycle arrest is independent of Mec1 in UV-irradiated Tel1-overproducing cells, while it is strictly Mec1 dependent in similarly treated DDC2-overexpressing cells. The Rad53 checkpoint kinase is instead required in both cases for cell cycle arrest, which correlates with its enhanced and persistent phosphorylation, suggesting that unscheduled Rad53 phosphorylation might prevent cells from re-entering the cell cycle after checkpoint activation. In addition, Tel1 overproduction results in transient nuclear division arrest and concomitant Rad53 phosphorylation in the absence of exogenous DNA damage independently of Mec1 and Ddc1.
Figures
References
-
- Chabes A., Domkin,V. and Thelander,L. (1999) Yeast Sml1, a protein inhibitor of ribonucleotide reductase. J. Biol. Chem., 274, 36679–36683. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
