

PepT1-mediated epithelial transport of dipeptides and cephalexin is enhanced by luminal leptin in the small intestine
- PMID: 11714740
- PMCID: PMC209419
- DOI: 10.1172/JCI13219
PepT1-mediated epithelial transport of dipeptides and cephalexin is enhanced by luminal leptin in the small intestine
Abstract
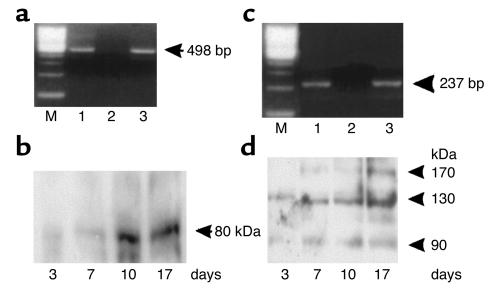
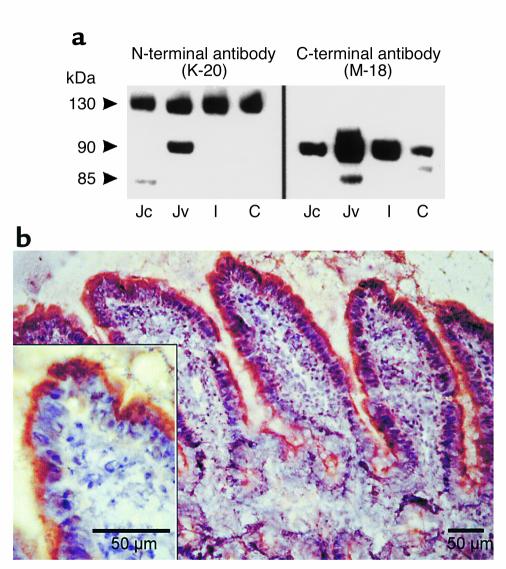
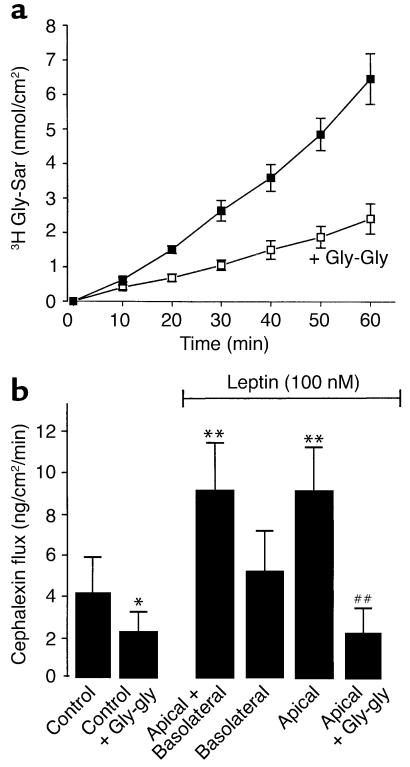
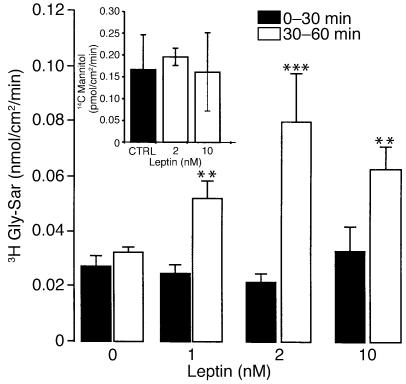
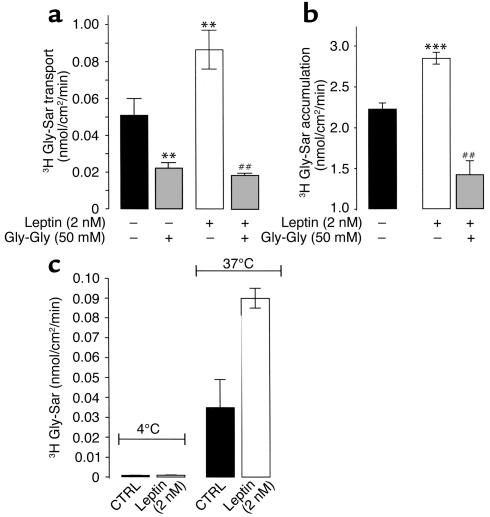
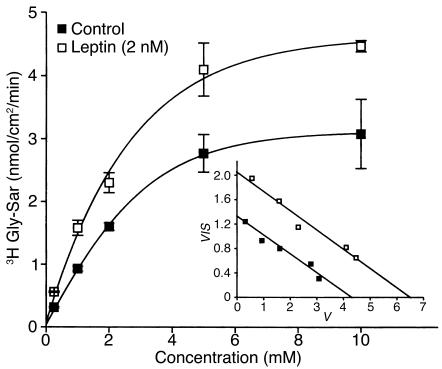
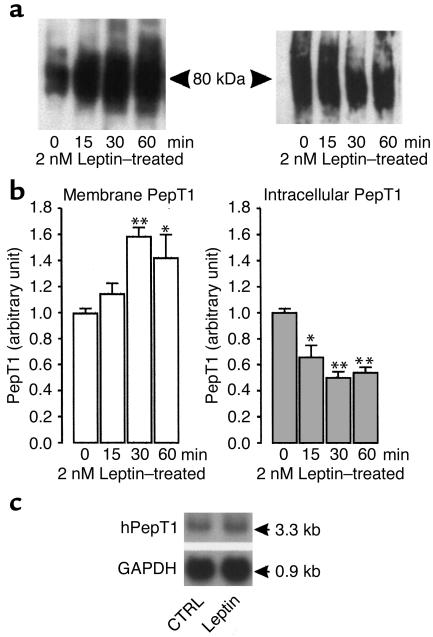
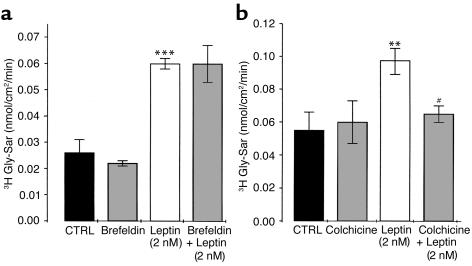
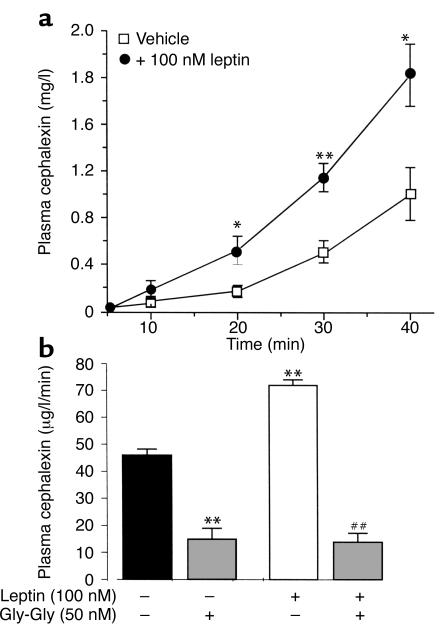
Dietary proteins are mostly absorbed as di- and tripeptides by the intestinal proton-dependent transporter PepT1. We have examined the effects of leptin on PepT1 function in rat jejunum and in monolayers of the human enterocyte-like 2 cell Caco-2. Leptin is produced by the stomach and secreted in the gut lumen. We show here that PepT1 and leptin receptors are expressed in Caco-2 and rat intestinal mucosal cells. Apical (but not basolateral) leptin increased Caco-2 cell transport of cephalexin (CFX) and glycylsarcosine (Gly-Sar), an effect that was associated with increased Gly-Sar uptake, increased membrane PepT1 protein, decreased intracellular PepT1 content, and no change in PepT1 mRNA levels. The maximal velocity (Vmax) for Gly-Sar transport was significantly increased by leptin, whereas the apparent Michaelis-Menten constant (Km) did not change. Furthermore, leptin-stimulated Gly-Sar transport was completely suppressed by colchicine, which disrupts cellular translocation of proteins to plasma membranes. Intrajejunal leptin also induced a rapid twofold increase in plasma CFX after jejunal perfusion with CFX in the rat, indicating enhanced intestinal absorption of CFX. These data revealed an unexpected action of gastric leptin in controlling ingestion of dietary proteins.
Figures
References
-
- Grimble GK. The significance of peptides in clinical nutrition. Annu Rev Nutr. 1994;14:419–447. - PubMed
-
- Adibi SA. The oligopeptide transporter (Pept-1) in human intestine: biology and function. Gastroenterology. 1997;113:332–340. - PubMed
-
- Liang R, et al. Human intestinal H+/peptide cotransporter. Cloning, functional expression, and chromosomal localization. J Biol Chem. 1995;270:6456–6463. - PubMed
-
- Boll M, et al. Expression cloning of a cDNA from rabbit small intestine related to proton-coupled transport of peptides, beta-lactam antibiotics and ACE-inhibitors. Pflugers Arch. 1994;429:146–149. - PubMed
-
- Fei YJ, et al. Expression cloning of a mammalian proton-coupled oligopeptide transporter. Nature. 1994;368:563–566. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
