

MxiM and MxiJ, base elements of the Mxi-Spa type III secretion system of Shigella, interact with and stabilize the MxiD secretin in the cell envelope
- PMID: 11717255
- PMCID: PMC95545
- DOI: 10.1128/JB.183.24.6991-6998.2001
MxiM and MxiJ, base elements of the Mxi-Spa type III secretion system of Shigella, interact with and stabilize the MxiD secretin in the cell envelope
Abstract
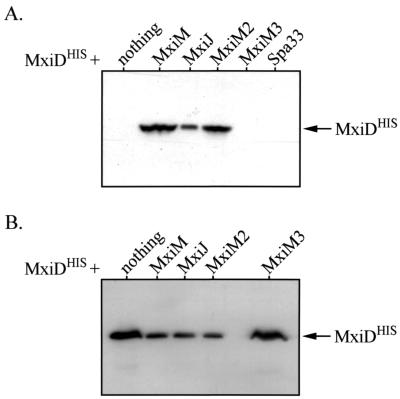
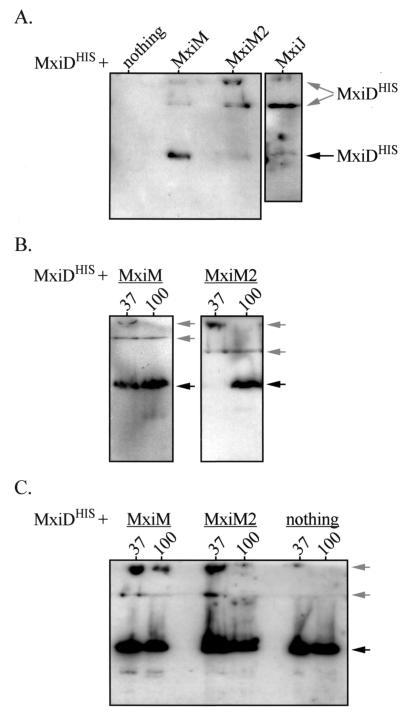
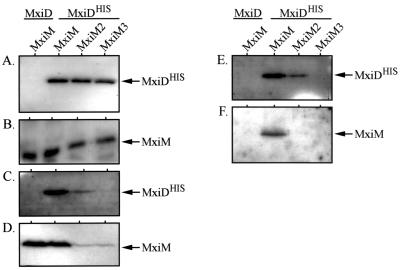
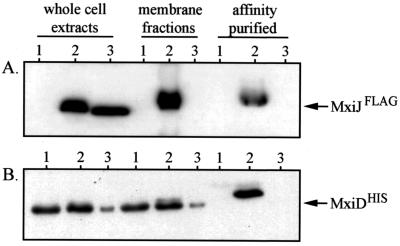
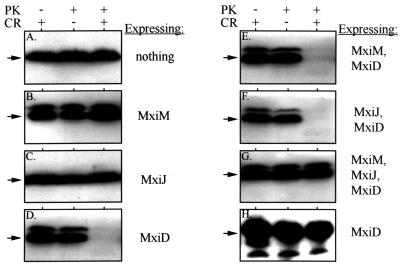
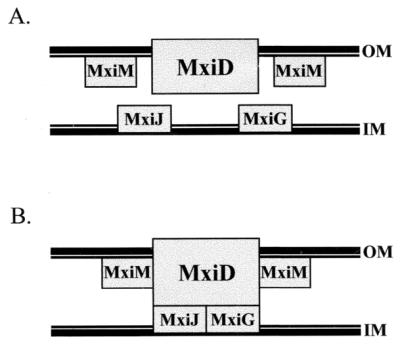
The type III secretion pathway is broadly distributed across many parasitic bacterial genera and serves as a mechanism for delivering effector proteins to eukaryotic cell surface and cytosolic targets. While the effectors, as well as the host responses elicited, differ among type III systems, they all utilize a conserved set of 9 to 11 proteins that together form a bacterial envelope-associated secretory organelle or needle complex. The general structure of the needle complex consists of a transenvelope base containing at least three ring-forming proteins (MxiD, MxiJ, and MxiG in Shigella) that is connected to a hollow needle-like extension that projects away from the cell surface. Several studies have shown that the initial steps in needle complex assembly require interactions among the base proteins, although specific details of this process remain unknown. Here we identify a role for another base element in Shigella, MxiM, in interactions with the major outer-membrane-associated ring-forming protein, MxiD. MxiM affects several features of MxiD, including its stability, envelope association, and assembly into homomultimeric structures. Interestingly, many of the effects were also elicited by the inner-membrane-associated base element, MxiJ. We confirmed that MxiM-MxiD and MxiJ-MxiD interactions occur in vivo in the cell envelope, and we present evidence that together these base elements can form a transmembrane structure which is likely an important intermediary in the process of needle complex assembly.
Figures
References
-
- Allaoui A, Sansonetti P J, Ménard R, Barzu S, Mounier J, Phalipon A, Parsot C. MxiG, a membrane protein required for secretion of Shigella spp. Ipa invasins: involvement in entry into epithelial cells and in intercellular dissemination. Mol Microbiol. 1995;17:461–470. - PubMed
-
- Allaoui A, Sansonetti P J, Parsot C. MxiD: an outer membrane protein necessary for the secretion of the Shigella flexneri Ipa invasins. Mol Microbiol. 1993;7:59–68. - PubMed
-
- Blocker A, Jouihri N, Larquet E, Gounon P, Ebel F, Parsot C, Sansonetti P, Allaoui A. Structure and composition of the Shigella flexneri ‘needle complex,’ a part of its type III secreton. Mol Microbiol. 2001;39:652–663. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
