

Review
doi: 10.1128/AEM.67.12.5351-5361.2001.
Microcompartments in prokaryotes: carboxysomes and related polyhedra
Affiliations
- PMID: 11722879
- PMCID: PMC93316
- DOI: 10.1128/AEM.67.12.5351-5361.2001
Item in Clipboard
Review
Microcompartments in prokaryotes: carboxysomes and related polyhedra
Appl Environ Microbiol.
2001 Dec.
No abstract available
Figures
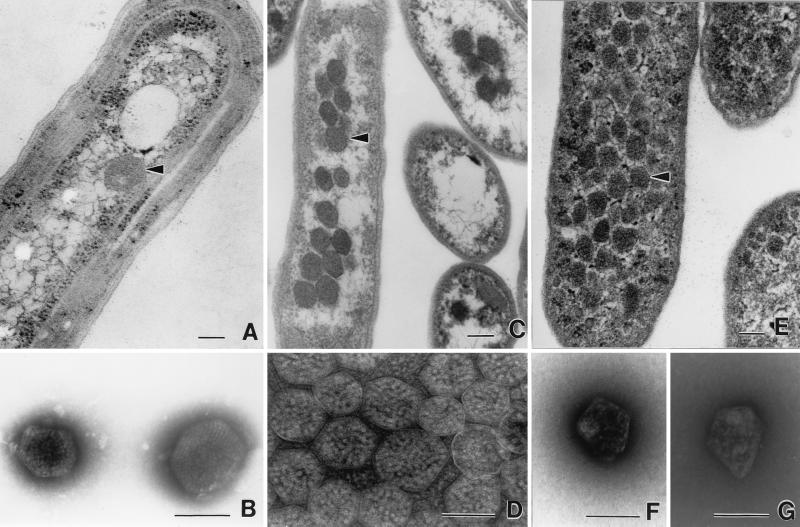
Transmission electron micrographs of carboxysomes and enterosomes. (A) Thin section of a cell of Synechococcus strain PCC7942 (fixed cells kindly supplied by George Espie), showing a typical carboxysome (arrowhead). (B) Negatively stained carboxysomes from lysed cells of A. nidulans (now Synechococcus). Molecules of RuBisCO are visible inside. Micrograph kindly supplied by Elisabeth Gantt. (C) Thin sections of H. neapolitanus grown in air, showing aggregation of carboxysomes (arrowhead) in the nucleoid region of the cell. (D) Negative stain of carboxysomes isolated from H. neapolitanus. RuBisCO assemblies are visible inside. (E) Thin section of S. enterica serovar Typhimurium LT2 grown on propanediol under aerobic conditions. Many polyhedral bodies (enterosomes [arrowhead]) are visible throughout the cytoplasm. They are less regular than carboxysomes and slightly smaller. (F and G) Negatively stained, isolated enterosomes from S. enterica serovar Typhimurium LT2. Note the irregular shape. Contents appear to be of variable sizes. Photographed from preparation kindly supplied by Greg Havemann. Panels A, C, and E are all printed at the same magnification, as are panels B, D, F, and G. Bars, 100 nm.
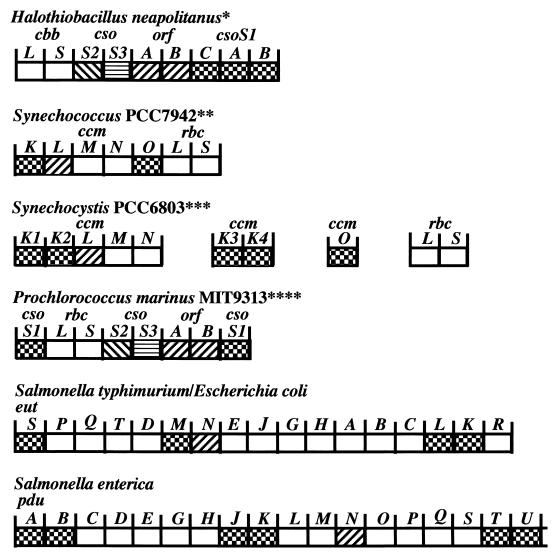
Organization of carboxysome genes and the genes of related polyhedra. Gene similarity is indicated by the different fill patterns. Gene homologies were assigned based on multiple sequence alignments that considered residue and positional identities. The deduced ORFA and ORFB peptides are 22 to 30% identical to CcmL gene products; CsoS1-like peptides are 50 to 99% identical (62). All of the genes are read from left to right. We have taken the liberty of calling the Prochlorococcus carboxysome genes csoS1, ORFAB, csoS2, and csoS3 instead of ccmK, ccmL, ccmM, and ccmN, respectively, since they are more closely related to the former. Abbreviations: cbb, Calvin-Benson-Bassham cycle in eubacteria and archaea cbbL and cbbS, large and small subunits of RuBisCO; cso, carboxysome; ORF, unidentified open reading frame; ccm, carbon-concentrating mechanism; rbcL and rbcS, large and small subunits of RuBisCO in cyanobacteria; pdu, propanediol utilization; eut, ethanolamine utilization. ∗, Two different sulfur bacteria exhibit the same organization, and a third exhibits the same except that the cso genes are not located downstream of cbbLS. ∗∗, Synechococcus strain PCC 7002 has the same organization as Synechococcus strain PCC7942, except that ccmO is missing from the cluster and rbcX resides between rbcL and rbcS. The ccmO gene may reside elsewhere on the genome of PCC7002 (see Synechocystis strain PCC6803). Additional ccmK genes could potentially reside elsewhere on the genome in both PCC7942 and PCC7002. Synechococcus strains PCC6301 possesses two ORFs immediately upstream of rbcL and rbcS, which appear to be equivalent to ccmO. Additional upstream sequencing has not been reported. ∗∗∗, Synechocystis strain PCC6803 possesses two additional ccmK genes and a ccmO gene completely separate from the ccmKLMN cluster. Nostoc punctiforme exhibits the same organization as Synechocystis strain PCC6803, except that ccmO is found immediately downstream of ccmN and not elsewhere on the genome. Furthermore, ccmO possess only a single ccmK homolog rather than two, as is the case with the other ccmO. ∗∗∗∗, Prochlorococcus strain MED4 shows the same organization as MIT9313, except the downstream csoS1 is missing. Synechococcus WH8102 shows the same organization as Prochlorococcus MIT9313. It seems likely that Synechococcus strain WH7803 will also show the same organization, but the downstream region has not been sequenced.
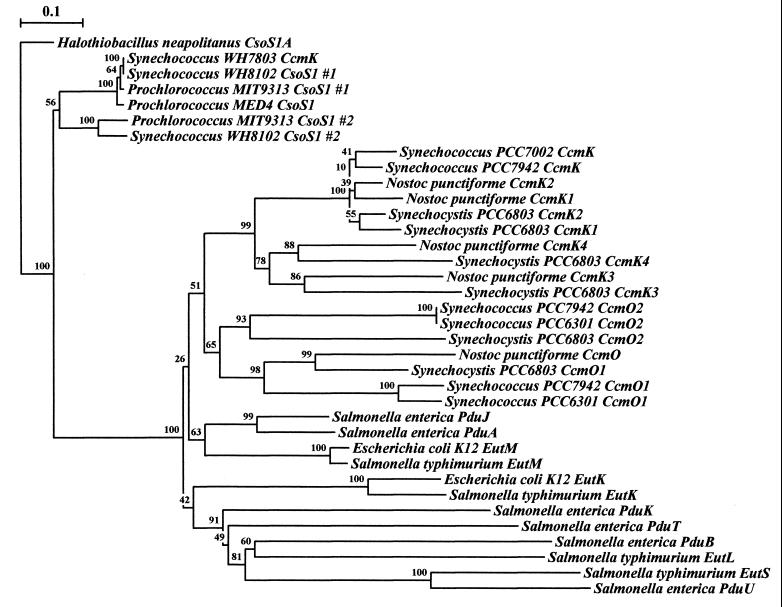
Phylogenetic tree of the conserved region of CsoS1-like polypeptides. Rooting was done arbitrarily using the conserved region of H. neapolitanus csoS1A, which codes for a polypeptide component of the carboxysome shell, which has greater than 90% similarity/identity to each of the known CsoS1 polypeptide regions. Several Synechococcus and Prochlorococcus species seem to group out near H. neapolitanus as well. Respective cyanobacterial and enteric CsoS1 homologs each seem to group together into respective major branches, with conserved protein clusters implying similarity of function and/or origin. The data for each bootstrap value were resampled 1,000 times, with the evolutionary distance scale (in the upper left-hand corner) horizontally representing the number of fixed substitutions per site. Sequence alignment was performed with ClustalW 1.8 (70), and phylogenetic trees were constructed with Treecon (72, 73).
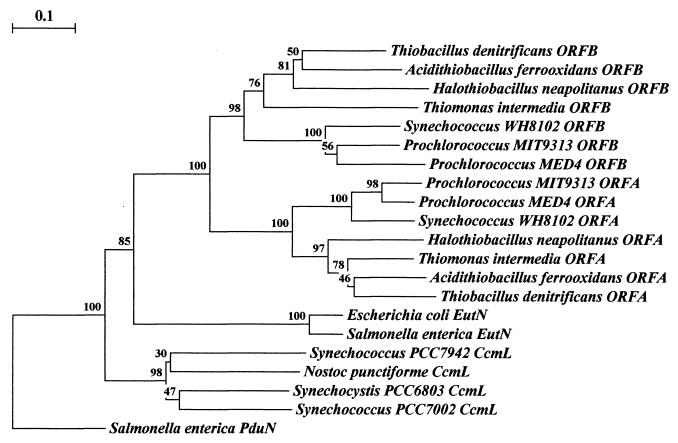
Phylogenetic tree of the ORFA and ORFB polypeptides and their relatives. Rooting was done arbitrarily through Salmonella enterica serovar Typhimurium PduN, which groups individually and away from the other families of polypeptides. Clustering the polypeptides into groupings respective of species physiology may be seen among the cyanobacteria, the thiobacilli, and the enteric polypeptides. Uniform clustering of protein homologs by relative physiology occurs with a high degree of confidence. Of the major branches of protein groupings, all are represented by bootstrap values greater than 75%. The data for each bootstrap value were resampled 1,000 times, with the evolutionary distance scale in the upper left-hand corner representing the number of fixed substitutions per site, as a measure of horizontal distance. Sequence alignment was performed with ClustalW 1.8 (70) and phylogenetic trees were constructed with Treecon (72, 73).
References
-
- Amichay D, Levitz R, Gurevitz M. Construction of a Synechocystis PCC6803 mutant suitable for the study of variant hexadecameric ribulose bisphosphate carboxylase/oxygenase enzymes. Plant Mol Biol. 1993;23:465–476. - PubMed
-
- Badger M R, Price G D. The CO2 concentrating mechanism in cyanobacteria and microalgae. Physiol Plant. 1992;90:529–536.
-
- Baker S H, Jin S, Aldrich H C, Howard G T, Shively J M. Insertion mutation of the form I cbbL gene encoding ribulose bisphosphate carboxylase/oxygenase (RuBisCO) in Thiobacillus neapolitanus results in expression of form II RuBisCO, loss of carboxysomes, and an increased CO2 2requirement for growth. J Bacteriol. 1998;180:4133–4139. - PMC - PubMed
-
- Baker S H, Lorbach S C, Rodriguez-Buey M, Williams D S, Aldrich H C, Shively J M. The correlation of the gene csoS2 of the carboxysome operon with two polypeptides of the carboxysome in Thiobacillus neapolitanus. Arch Microbiol. 1999;172:233–239. - PubMed
-
- Baker S H, Williams D S, Aldrich H C, Gambrell A C, Shively J M. Identification and localization of the carboxysome peptide Csos3 and its corresponding gene in Thiobacillus neapolitanus. Arch Microbiol. 2000;173:278–283. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
