

Specific interference with gene expression induced by long, double-stranded RNA in mouse embryonal teratocarcinoma cell lines
- PMID: 11724966
- PMCID: PMC64698
- DOI: 10.1073/pnas.261562698
Specific interference with gene expression induced by long, double-stranded RNA in mouse embryonal teratocarcinoma cell lines
Abstract
In eukaryotes, double-stranded (ds) RNA induces sequence-specific inhibition of gene expression, referred to as RNA interference (RNAi). In invertebrates, RNAi can be triggered effectively by either long dsRNAs or 21- to 23-nt-long short interfering (si) duplex RNAs, acting as effectors of RNAi. siRNAs recently have been shown to act as potent inducers of RNAi in cultured mammalian cells. However, studies of RNAi activated by long dsRNA are impeded by its nonspecific effects, mediated by dsRNA-dependent protein kinase PKR and RNase L. Here, we report that the RNAi response can be induced effectively by long dsRNA in nondifferentiated mouse cells grown in culture. Transfection of dsRNA into embryonal carcinoma (EC) P19 and F9 cells results in a sequence-specific decrease in the level of proteins expressed from either exogenous or endogenous genes. dsRNA-mediated inhibition of the reporter gene also occurs in mouse embryonic stem cells. The RNAi effect is mediated by siRNAs, which are generated by cleavage of dsRNA by the RNaseIII-like enzyme, Dicer. We demonstrate that extracts prepared from EC cells catalyze processing of dsRNA into approximately 23-nt fragments and that Dicer localizes to the cytoplasm of EC and HeLa cells.
Figures
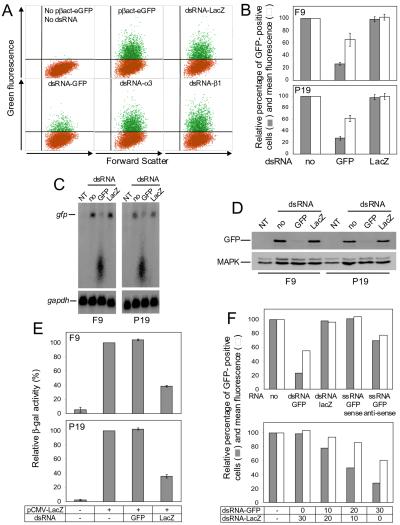
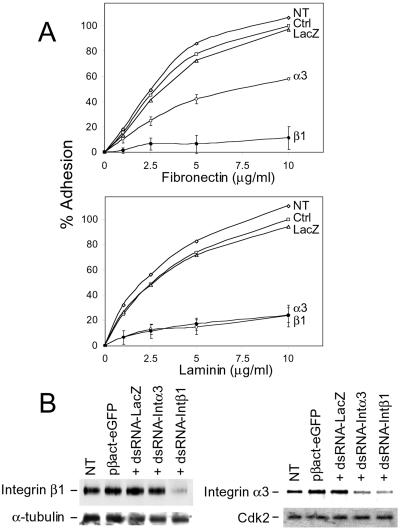
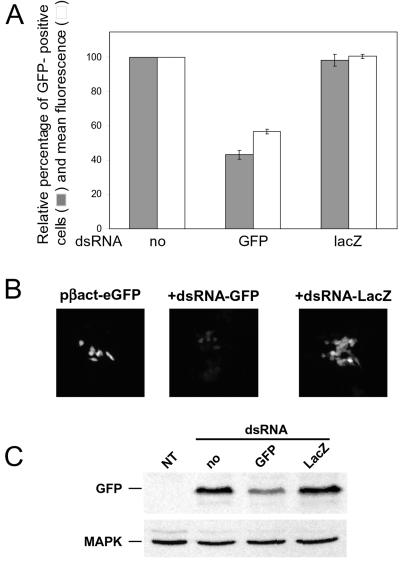
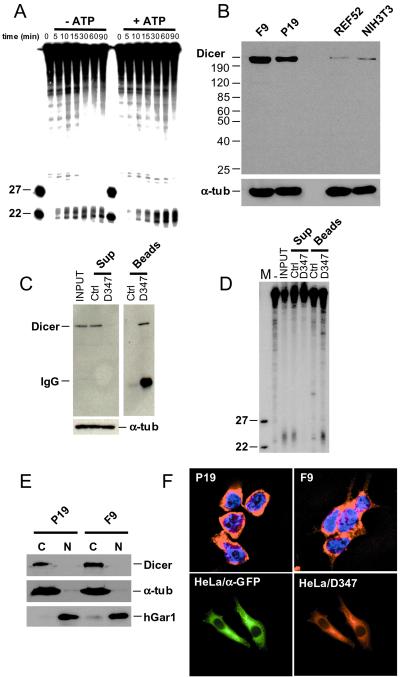
References
-
- Hammond S M, Caudy A A, Hannon G J. Nat Rev Genet. 2001;2:110–119. - PubMed
-
- Matzke M A, Matzke A J, Pruss G J, Vance V B. Curr Opin Genet Dev. 2001;11:221–227. - PubMed
-
- Sharp P A. Genes Dev. 2001;15:485–490. - PubMed
-
- Grishok A, Pasquinelli A E, Conte D, Li N, Parrish S, Ha I, Baillie D L, Fire A, Ruvkun G, Mello C C. Cell. 2001;106:23–34. - PubMed
-
- Hutvagner G, McLachlan J, Pasquinelli A E, Balint E, Tuschl T, Zamore P D. Science. 2001;293:834–838. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
