

Control of branch-site choice by a group II intron
- PMID: 11726522
- PMCID: PMC125754
- DOI: 10.1093/emboj/20.23.6866
Control of branch-site choice by a group II intron
Abstract
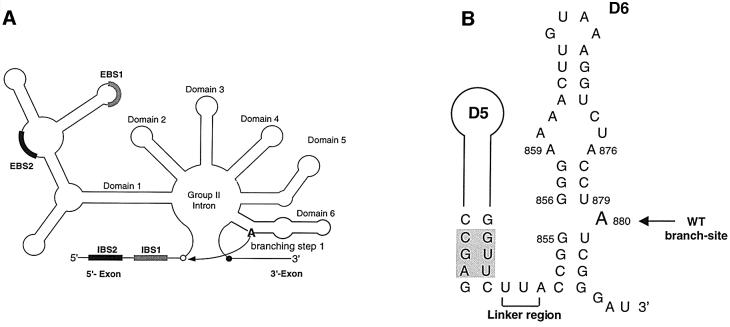
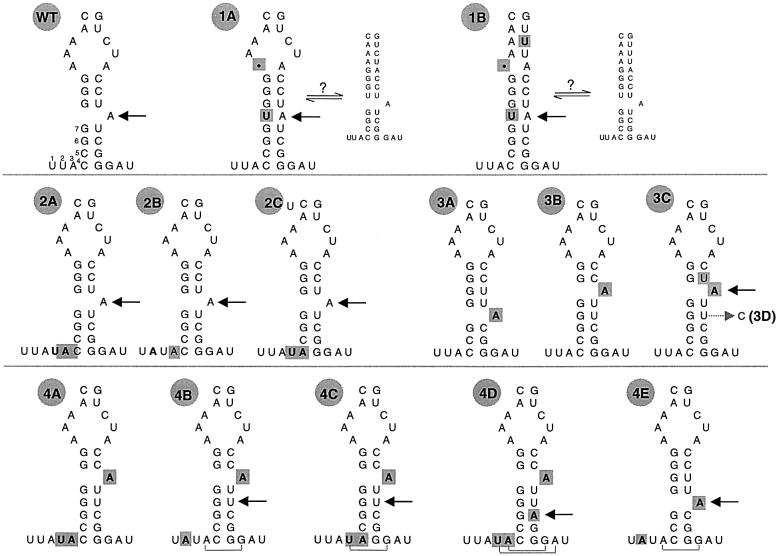
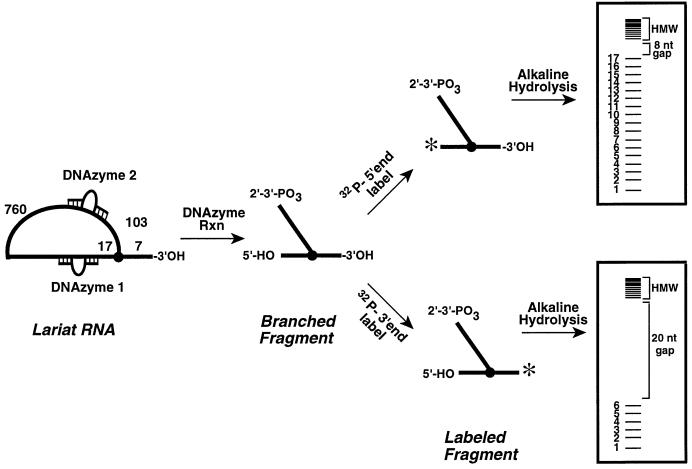
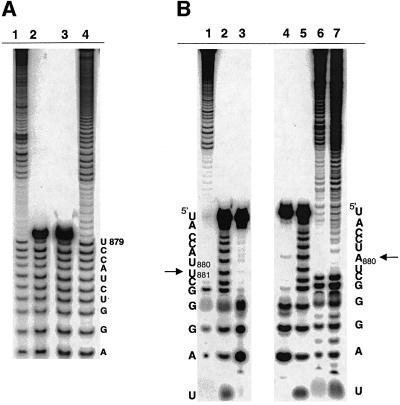
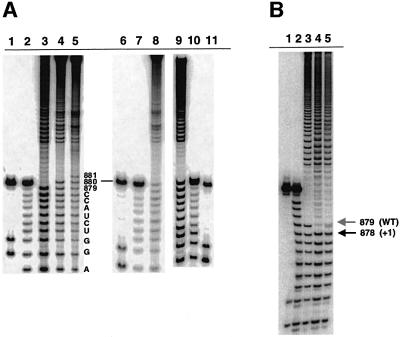
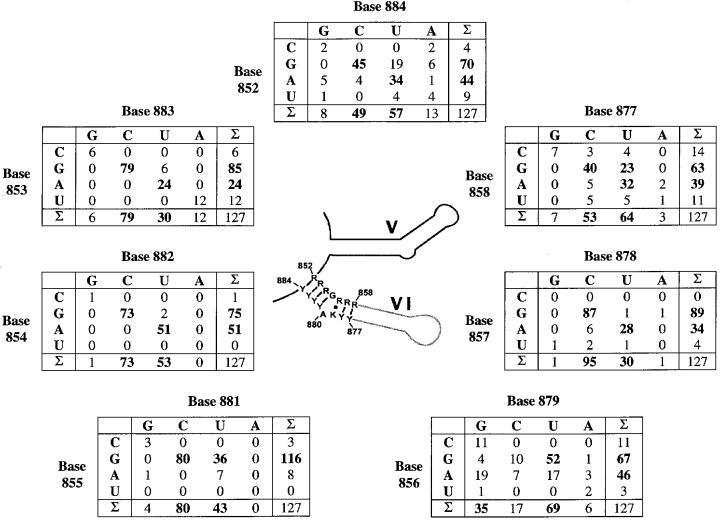
The branch site of group II introns is typically a bulged adenosine near the 3'-end of intron domain 6. The branch site is chosen with extraordinarily high fidelity, even when the adenosine is mutated to other bases or if the typically bulged adenosine is paired. Given these facts, it has been difficult to discern the mechanism by which the proper branch site is chosen. In order to dissect the determinants for branch-point recognition, new mutations were introduced in the vicinity of the branch site and surrounding domains. Single mutations did not alter the high fidelity for proper branch-site selection. However, several combinations of mutations moved the branch site systematically to new positions along the domain 6 stem. Analysis of those mutants, together with a new alignment of domain 5 and domain 6 sequences, reveals a set of structural determinants that appear to govern branch-site selection by group II introns.
Figures

References
-
- Bonen L. and Vogel,J. (2001) The ins and outs of group II introns. Trends Genet., 17, 322–331. - PubMed
-
- Burkard M.E., Kierzek,R. and Turner,D.H. (1999) Thermodynamics of unpaired terminal nucleotides on short RNA helices correlates with stacking at helix termini in larger RNAs. J. Mol. Biol., 290, 967–982. - PubMed
-
- Cech T.R. (1986) The generality of self-splicing RNA: relationship to nuclear mRNA splicing. Cell, 44, 207–210. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
