

Creatine kinase is physically associated with the cardiac ATP-sensitive K+ channel in vivo
- PMID: 11729098
- PMCID: PMC2130771
- DOI: 10.1096/fj.01-0466fje
Creatine kinase is physically associated with the cardiac ATP-sensitive K+ channel in vivo
Abstract
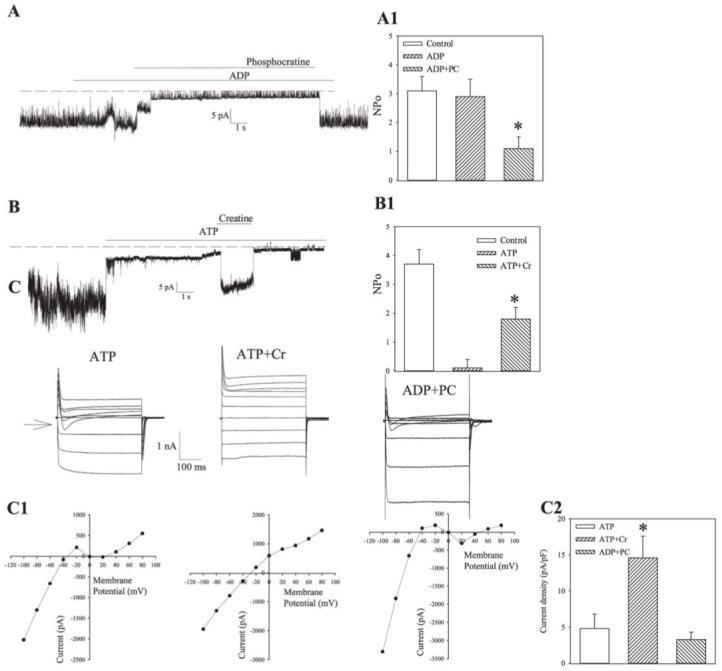
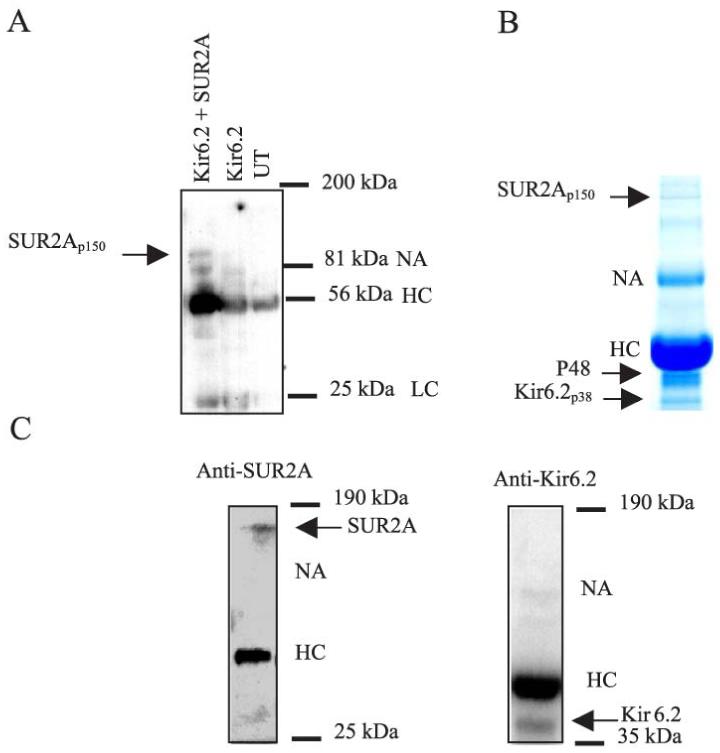
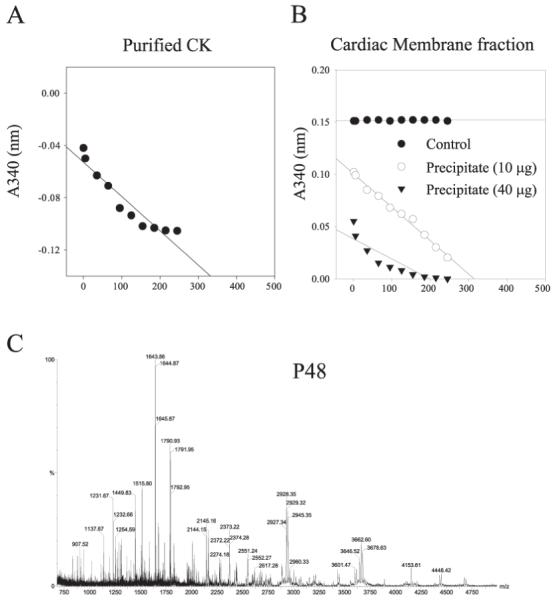
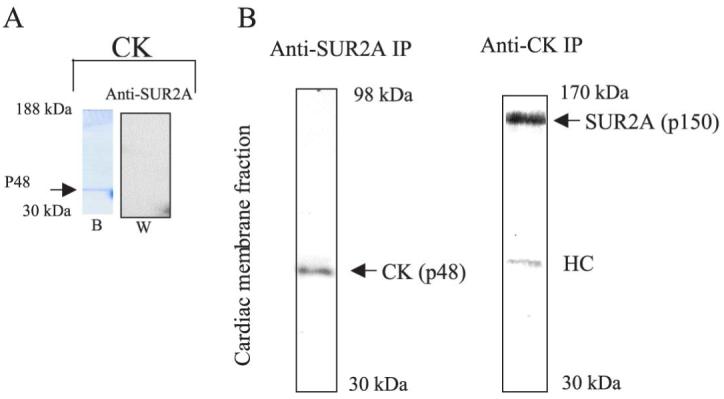
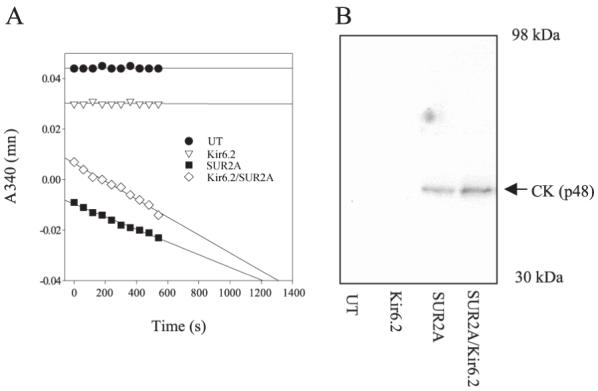
Cardiac sarcolemmal ATP-sensitive K+ (KATP) channels, composed of Kir6.2 and SUR2A subunits, couple the metabolic status of cells with the membrane excitability. Based on previous functional studies, we have hypothesized that creatine kinase (CK) may be a part of the sarcolemmal KATP channel protein complex. The inside-out and whole cell patch clamp electrophysiology applied on guinea pig cardiomyocytes showed that substrates of CK regulate KATP channels activity. Following immunoprecipitation of guinea-pig cardiac membrane fraction with the anti-SUR2 antibody, Coomassie blue staining revealed, besides Kir6.2 and SUR2A, a polypeptide at approximately 48 kDa. Western blotting analysis confirmed the nature of putative Kir6.2 and SUR2A, whereas matrix-assisted laser desorption/ionization time-of-flight mass spectrometry analysis identified p48 kDa as a muscle form of CK. In addition, the CK activity was found in the anti-SUR2A immunoprecipitate and the cross reactivity between an anti-CK antibody and the anti-SUR2A immunoprecipitate was observed as well as vice verse. Further results obtained at the level of recombinant channel subunits demonstrated that CK is directly physically associated with the SUR2A, but not the Kir6.2, subunit. All together, these results suggest that the CK is associated with SUR2A subunit in vivo, which is an integral part of the sarcolemmal KATP channel protein complex.
Figures
References
-
- Dzeja PP, Terzic A. Phosphotransfer reactions in the regulation of ATP-sensitive K+ channels. FASEB J. 1998;12:523–529. - PubMed
-
- Wyss M, Kaddurah-Daouk R. Creatine and creatinine metabolism. Physiol. Rev. 2000;80:1107–1213. - PubMed
-
- Noma A. ATP-regulated K+ channels in cardiac muscle. Nature (London) 1983;305:147–148. - PubMed
-
- Gross GJ, Fryer RM. Sarcolemmal versus mitochondrial ATP-sensitive K+ channels and myocardial preconditioning. Circ. Res. 1999;84:973–979. - PubMed
-
- Jovanovic A, Jovanovic S, Lorenz E, Terzic A. Recombinant cardiac ATP-sensitive K+ channel subunits confer resistance towards chemical hypoxia-reoxygenation injury. Circulation. 1998;98:1548–1555. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
