

Degradation of p53 by adenovirus E4orf6 and E1B55K proteins occurs via a novel mechanism involving a Cullin-containing complex
- PMID: 11731475
- PMCID: PMC312842
- DOI: 10.1101/gad.926401
Degradation of p53 by adenovirus E4orf6 and E1B55K proteins occurs via a novel mechanism involving a Cullin-containing complex
Abstract
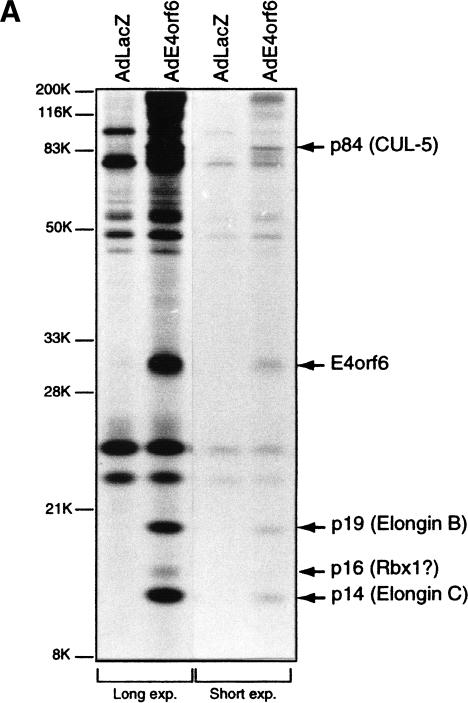
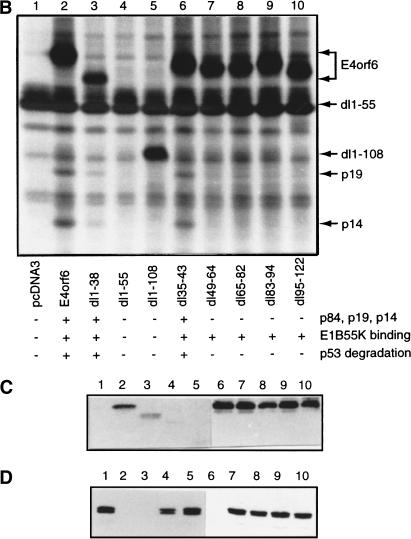
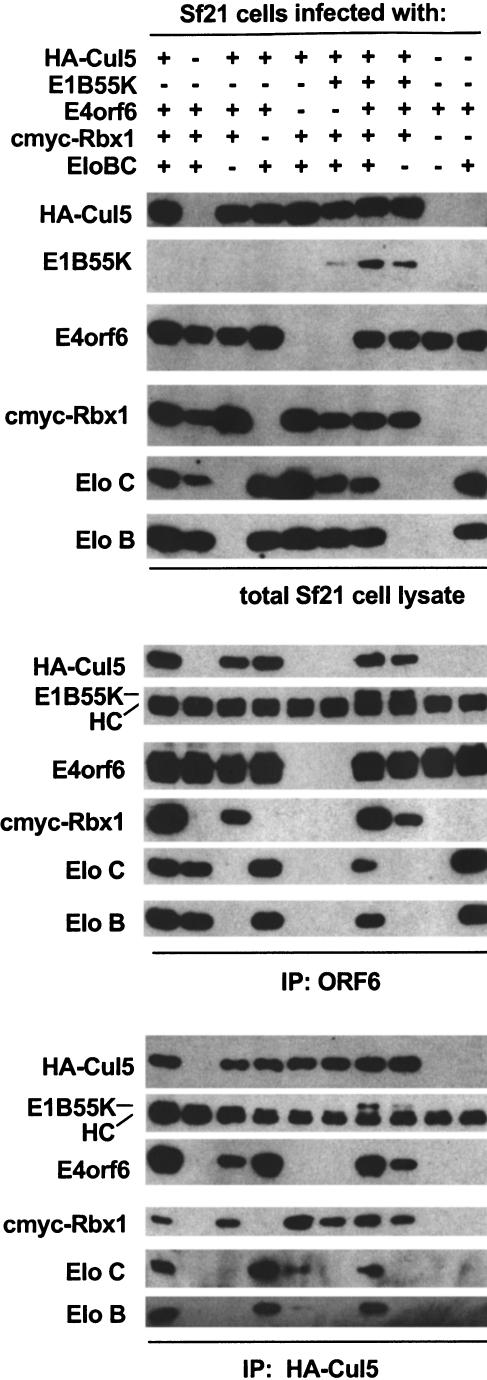
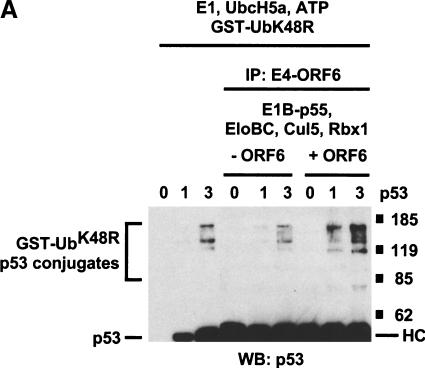
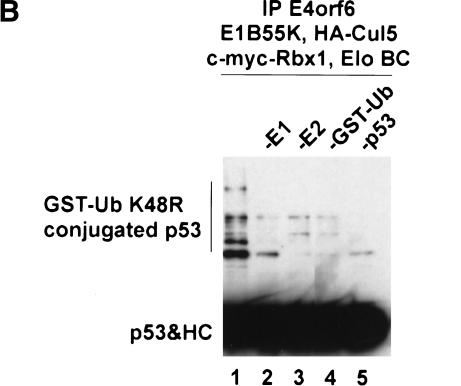
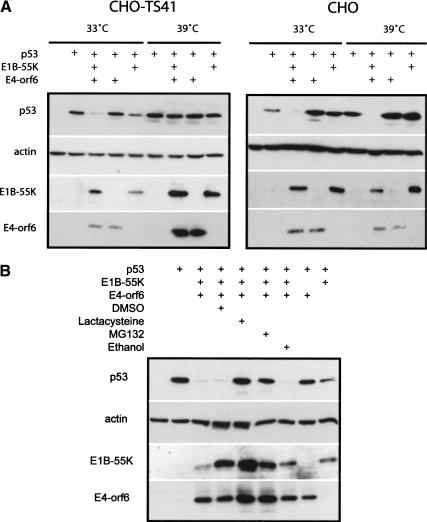
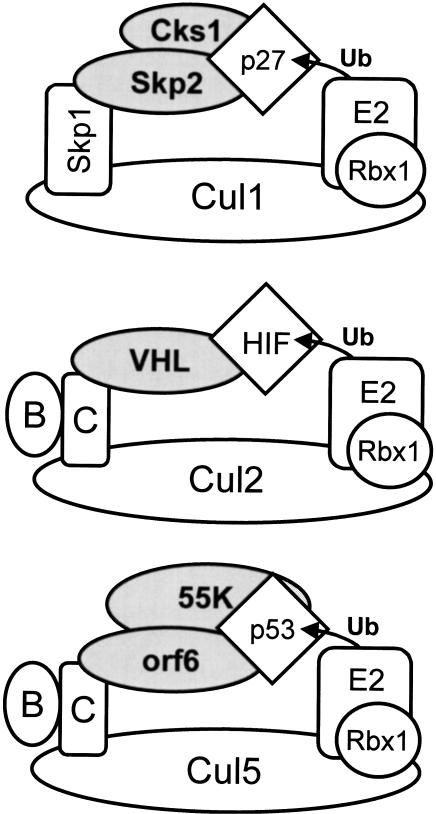
Although MDM2 plays a major role in regulating the stability of the p53 tumor suppressor protein, other poorly understood MDM2-independent pathways also exist. Human adenoviruses have evolved strategies to regulate p53 function and stability to permit efficient viral replication. One mechanism involves adenovirus E1B55K and E4orf6 proteins, which collaborate to target p53 for degradation. To determine the mechanism of this process, a multiprotein E4orf6-associated complex was purified and shown to contain a novel Cullin-containing E3 ubiquitin ligase that is (1) composed of Cullin family member Cul5, Elongins B and C, and the RING-H2 finger protein Rbx1(ROC1); (2) remarkably similar to the von Hippel-Lindau tumor suppressor and SCF (Skp1-Cul1/Cdc53-F-box) E3 ubiquitin ligase complexes; and (3) capable of stimulating ubiquitination of p53 in vitro in the presence of E1/E2 ubiquitin-activating and -conjugating enzymes. Cullins are activated by NEDD8 modification; therefore, to determine whether Cullin complexes are required for adenovirus-induced p53 degradation, studies were conducted in ts41 Chinese hamster ovary cells that are temperature sensitive for the NEDD8 pathway. E4orf6/E1B55K failed to induce the degradation of p53 at the nonpermissive temperature. Thus, our results identify a novel role for the Cullin-based machinery in regulation of p53.
Figures

References
-
- Bacchetti S, Graham FL. Inhibition of cell proliferation by an adenovirus vector expressing the human wild type p53 protein. Int J Oncol. 1993;3:781–788. - PubMed
-
- Bargonetti J, Reynisdottir I, Friedman PN, Prives C. Site-specific binding of wild-type p53 to cellular DNA is inhibited by SV40 T antigen and mutant p53. Genes & Dev. 1992;6:1886–1898. - PubMed
-
- Bartek J, Lukas J. p27 destruction: Cks1 pulls the trigger. Nat Cell Biol. 2001;3:E95–E98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous