

Histone H3 specific acetyltransferases are essential for cell cycle progression
- PMID: 11731478
- PMCID: PMC312843
- DOI: 10.1101/gad.931401
Histone H3 specific acetyltransferases are essential for cell cycle progression
Abstract
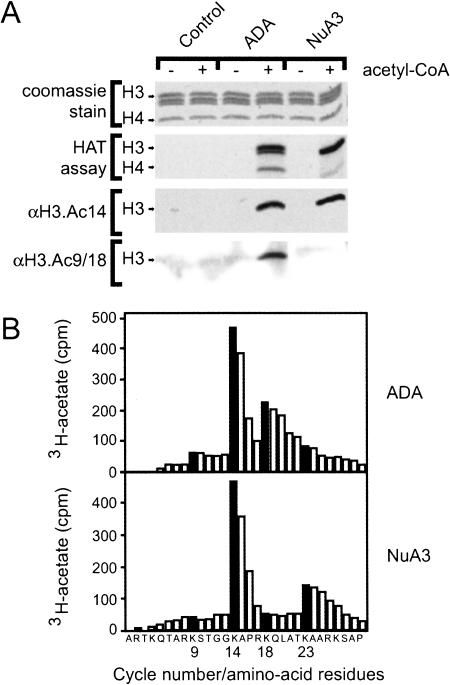
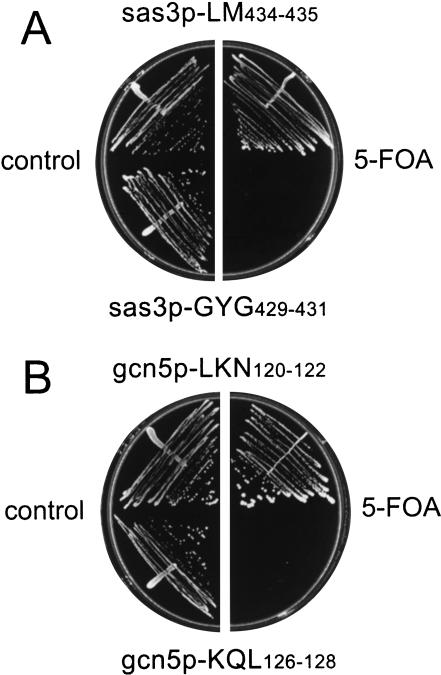
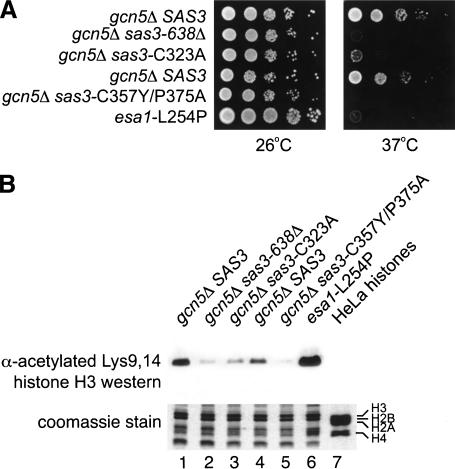
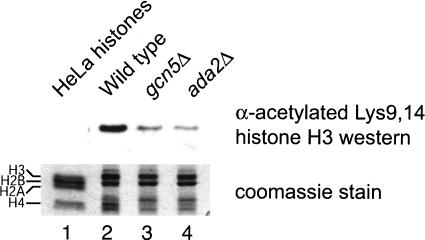
Longstanding observations suggest that acetylation and/or amino-terminal tail structure of histones H3 and H4 are critical for eukaryotic cells. For Saccharomyces cerevisiae, loss of a single H4-specific histone acetyltransferase (HAT), Esa1p, results in cell cycle defects and death. In contrast, although several yeast HAT complexes preferentially acetylate histone H3, the catalytic subunits of these complexes are not essential for viability. To resolve the apparent paradox between the significance of H3 versus H4 acetylation, we tested the hypothesis that H3 modification is essential, but is accomplished through combined activities of two enzymes. We observed that Sas3p and Gcn5p HAT complexes have overlapping patterns of acetylation. Simultaneous disruption of SAS3, the homolog of the MOZ leukemia gene, and GCN5, the hGCN5/PCAF homolog, is synthetically lethal due to loss of acetyltransferase activity. This key combination of activities is specific for these two HATs because neither is synthetically lethal with mutations of other MYST family or H3-specific acetyltransferases. Further, the combined loss of GCN5 and SAS3 functions results in an extensive, global loss of H3 acetylation and arrest in the G(2)/M phase of the cell cycle. The strikingly similar effect of loss of combined essential H3 HAT activities and the loss of a single essential H4 HAT underscores the fundamental biological significance of each of these chromatin-modifying activities.
Figures

References
-
- Adams A, Gottschling DE, Kaiser CA, Stearns T. Methods in yeast genetics: A Cold Spring Harbor Laboratory course manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1997.
-
- Angus-Hill ML, Dutnall RN, Tafrov ST, Sternglanz R, Ramakrishnan V. Crystal structure of the histone acetyltransferase Hpa2: A tetrameric member of the Gcn5-related N-acetyltransferase superfamily. J Mol Biol. 1999;294:1311–1325. - PubMed
-
- Ausubel FM. Current protocols in molecular biology. NY: Greene Pub. Associates and Wiley-Interscience, J. Wiley; 1987.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases