

Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications
- PMID: 11731568
- PMCID: PMC2278966
- DOI: 10.1111/j.1469-7793.2001.00333.x
Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications
Abstract
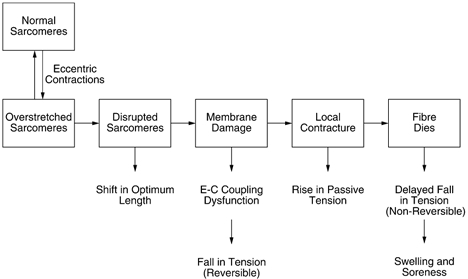
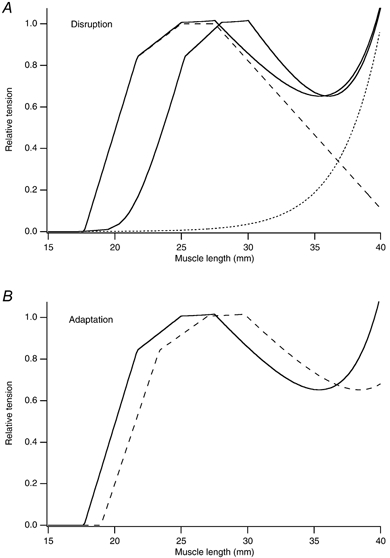
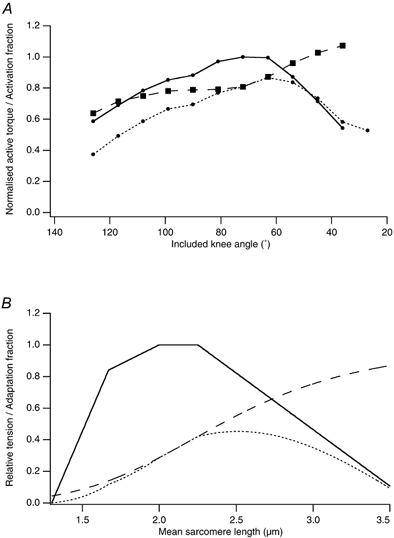
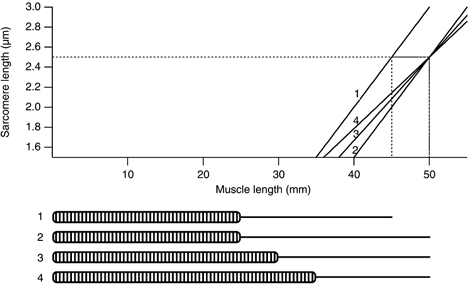
In eccentric exercise the contracting muscle is forcibly lengthened; in concentric exercise it shortens. While concentric contractions initiate movements, eccentric contractions slow or stop them. A unique feature of eccentric exercise is that untrained subjects become stiff and sore the day afterwards because of damage to muscle fibres. This review considers two possible initial events as responsible for the subsequent damage, damage to the excitation-contraction coupling system and disruption at the level of the sarcomeres. Other changes seen after eccentric exercise, a fall in active tension, shift in optimum length for active tension, and rise in passive tension, are seen, on balance, to favour sarcomere disruption as the starting point for the damage. As well as damage to muscle fibres there is evidence of disturbance of muscle sense organs and of proprioception. A second period of exercise, a week after the first, produces much less damage. This is the result of an adaptation process. One proposed mechanism for the adaptation is an increase in sarcomere number in muscle fibres. This leads to a secondary shift in the muscle's optimum length for active tension. The ability of muscle to rapidly adapt following the damage from eccentric exercise raises the possibility of clinical applications of mild eccentric exercise, such as for protecting a muscle against more major injuries.
Figures
References
-
- Allen DG. Eccentric muscle damage: mechanisms of early reduction of force. Acta Physiologica Scandanavica. 2001;171:311–319. - PubMed
-
- Allen TA. PhD Thesis. Clayton, Victoria, Australia: Monash University; 1999. Effect of eccentric contractions on the mechanical properties of skeletal muscle.
-
- Appell HJ, Soares JM, Duarte JA. Exercise, muscle damage and fatigue. Journal of Sports Medicine. 1992;13:108–115. - PubMed
-
- Armstrong RB, Ogilvie RW, Schwane JA. Eccentric exercise-induced injury to rat skeletal muscle. Journal of Applied Physiology. 1983;54:80–93. - PubMed
-
- Armstrong RB, Warren GL, Warren JA. Mechanisms of exercise-induced muscle fibre injury. Journal of Sports Medicine. 1991;12:184–207. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials