

Selective insolubility of alpha-synuclein in human Lewy body diseases is recapitulated in a transgenic mouse model
- PMID: 11733371
- PMCID: PMC1850592
- DOI: 10.1016/s0002-9440(10)63072-6
Selective insolubility of alpha-synuclein in human Lewy body diseases is recapitulated in a transgenic mouse model
Abstract
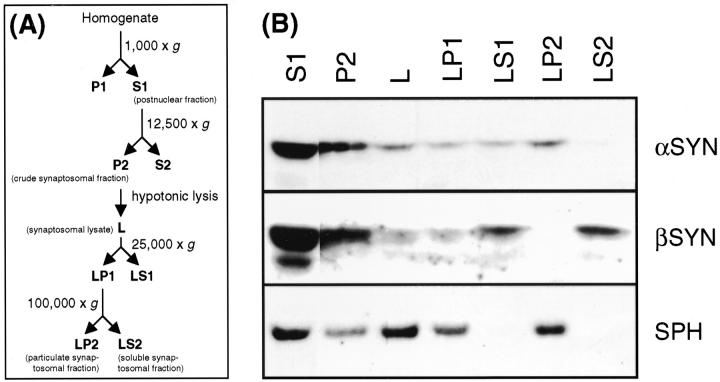
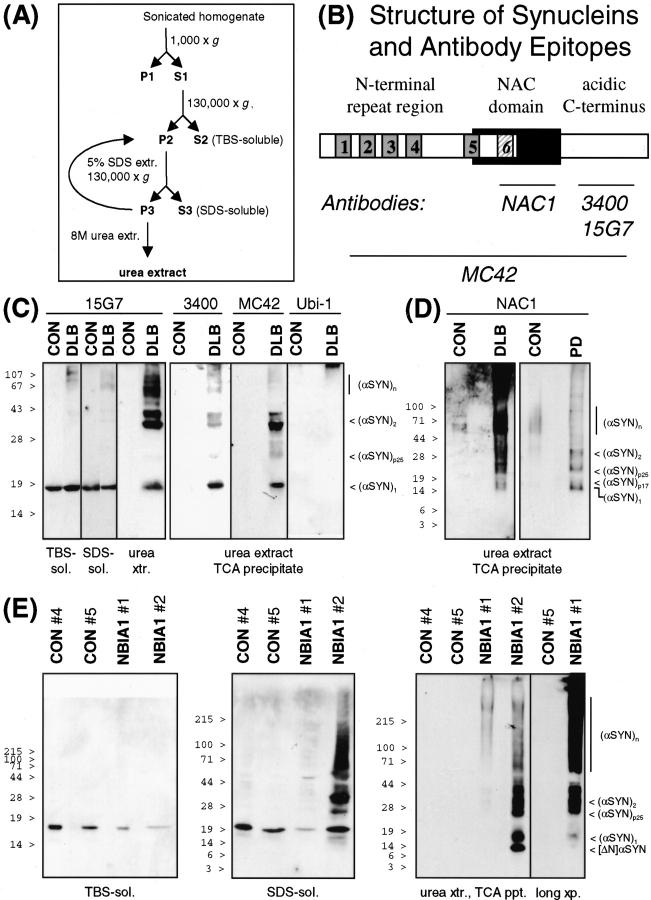
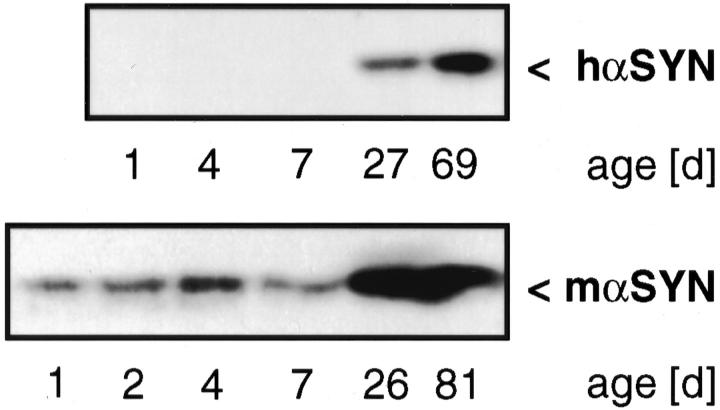
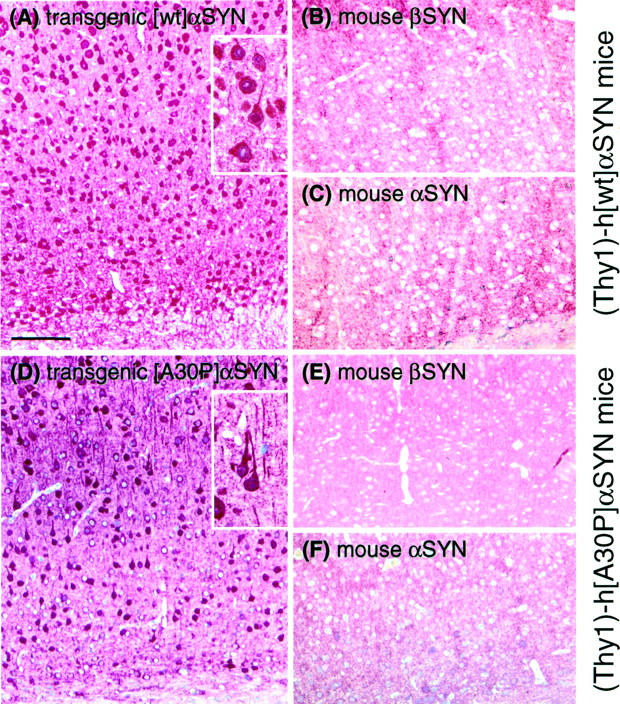
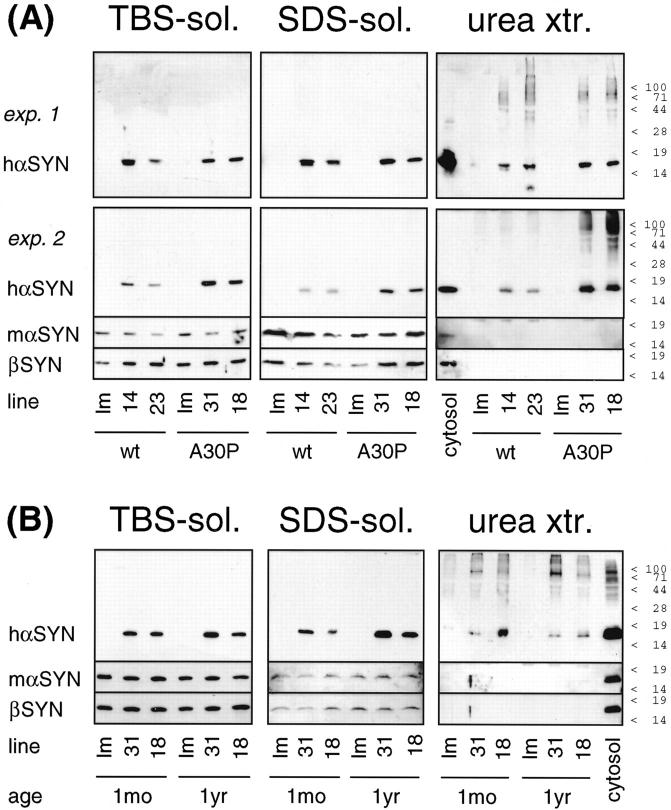
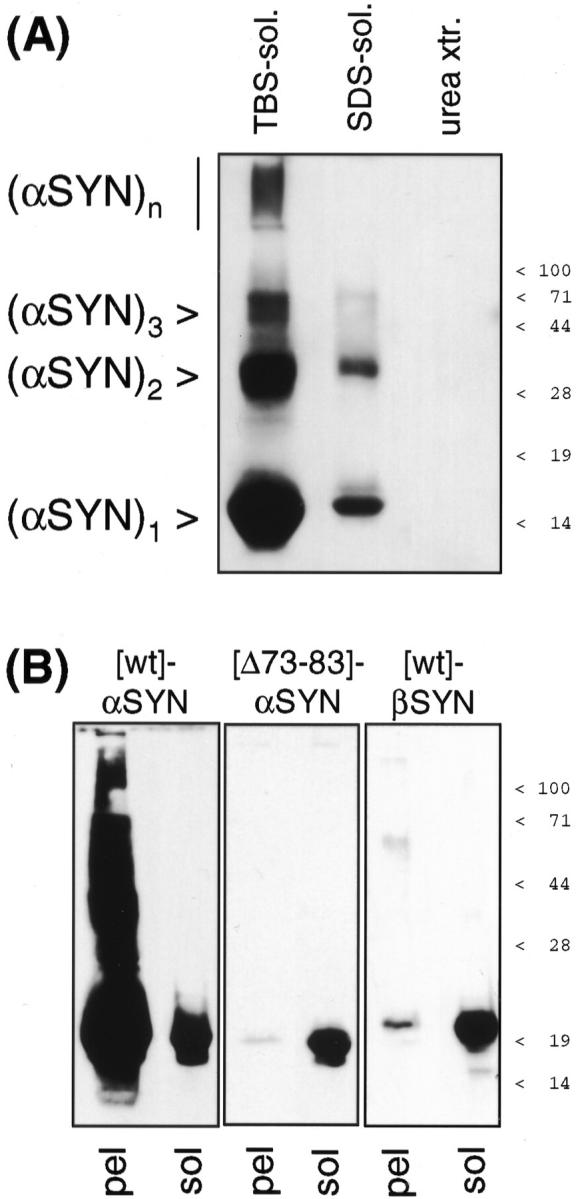
alpha-Synuclein (alpha-SYN) is deposited in intraneuronal cytoplasmic inclusions (Lewy bodies, LBs) characteristic for Parkinson's disease (PD) and LB dementias. alpha-SYN forms LB-like fibrils in vitro, in contrast to its homologue beta-SYN. Here we have investigated the solubility of SYNs in human LB diseases and in transgenic mice expressing human wild-type and PD-associated mutant [A30P]alpha-SYN driven by the brain neuron-specific promoter, Thy1. Distinct alpha-SYN species were detected in the detergent-insoluble fractions from brains of patients with PD, dementia with LBs, and neurodegeneration with brain iron accumulation type 1 (formerly known as Hallervorden-Spatz disease). Using the same extraction method, detergent-insolubility of human alpha-SYN was observed in brains of transgenic mice. In contrast, neither endogenous mouse alpha-SYN nor beta-SYN were detected in detergent-insoluble fractions from transgenic mouse brains. The nonamyloidogenic beta-SYN was incapable of forming insoluble fibrils because amino acids 73 to 83 in the central region of alpha-SYN are absent in beta-SYN. In conclusion, the specific accumulation of detergent-insoluble alpha-SYN in transgenic mice recapitulates a pivotal feature of human LB diseases.
Figures
References
-
- Spillantini MG, Schmidt ML, Lee VM-Y, Trojanowski JQ, Jakes R, Goedert M: α-Synuclein in Lewy bodies. Nature 1997, 388:839-840 - PubMed
-
- Wakabayashi K, Matsumoto K, Takayama K, Yoshimoto M, Takahashi H: NACP, a presynaptic protein, immunoreactivity in Lewy bodies in Parkinson’s disease. Neurosci Lett 1997, 239:45-48 - PubMed
-
- Irizarry MC, Growdon W, Gomez-Isla T, Newell K, George JM, Clayton DF, Hyman BT: Nigral and cortical Lewy bodies and dystrophic nigral neurites in Parkinson’s disease and cortical Lewy body disease contain α-synuclein immunoreactivity. J Neuropathol Exp Neurol 1998, 57:334-337 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
