

Thin filaments elongate from their pointed ends during myofibril assembly in Drosophila indirect flight muscle
- PMID: 11739412
- PMCID: PMC2150893
- DOI: 10.1083/jcb.200108026
Thin filaments elongate from their pointed ends during myofibril assembly in Drosophila indirect flight muscle
Abstract
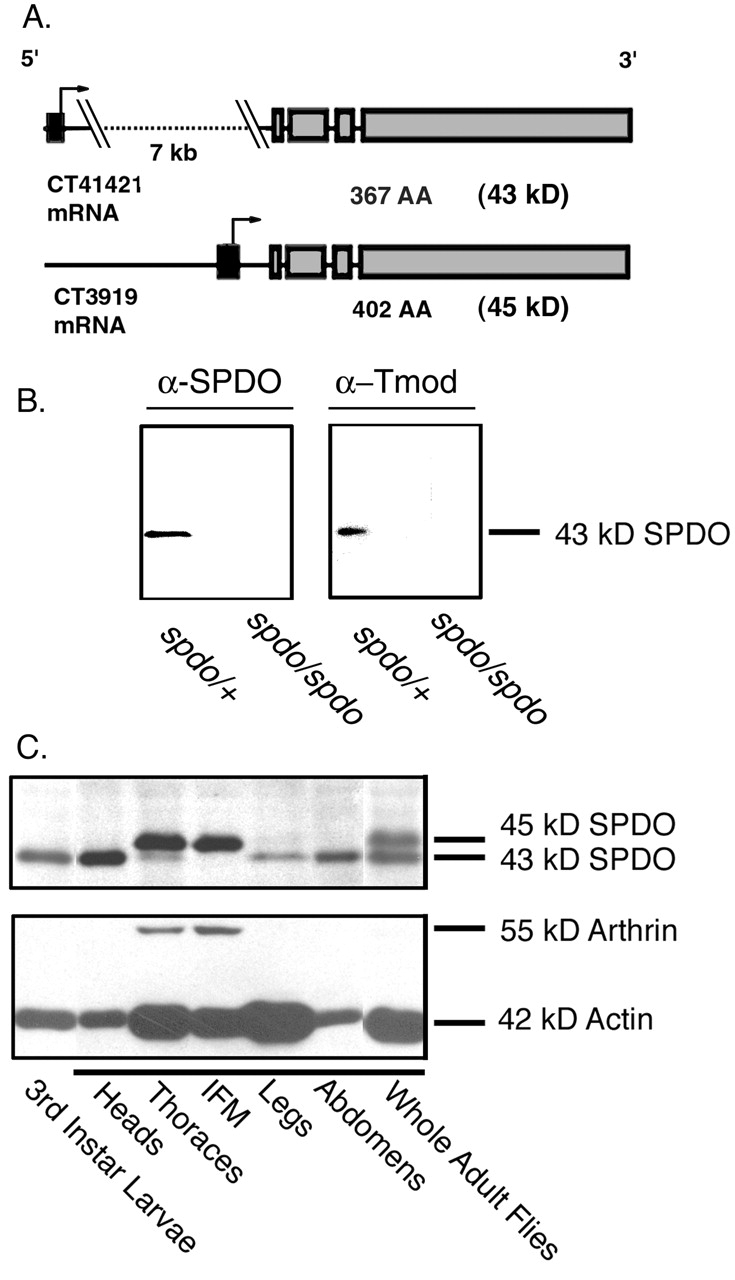
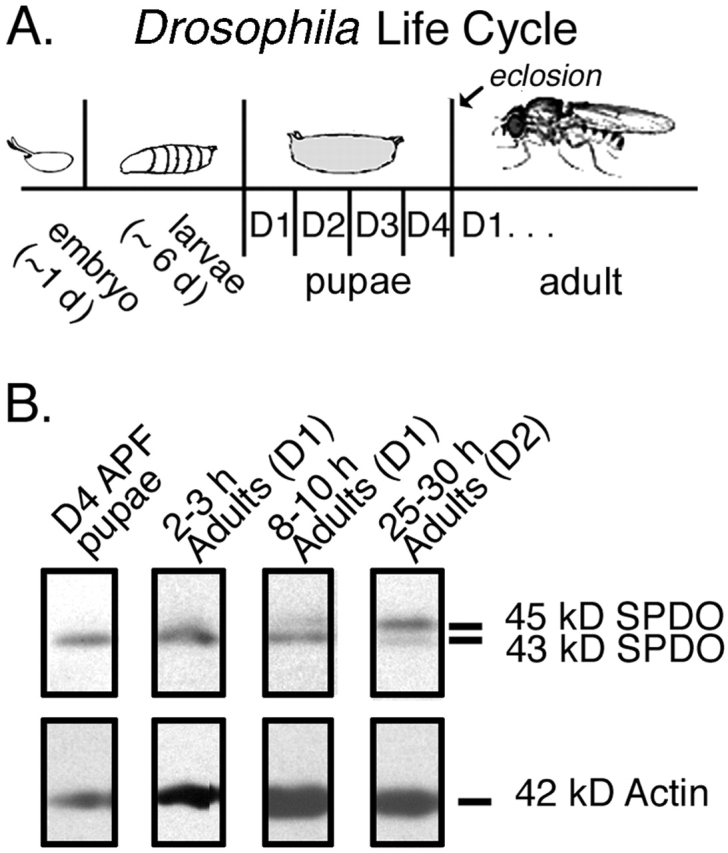
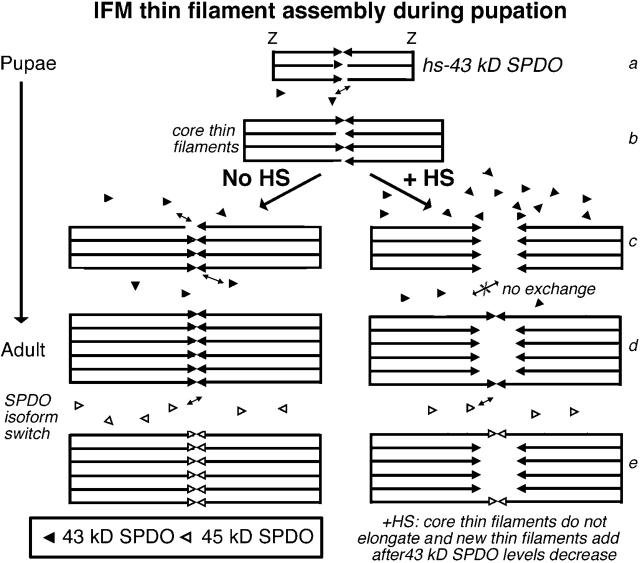
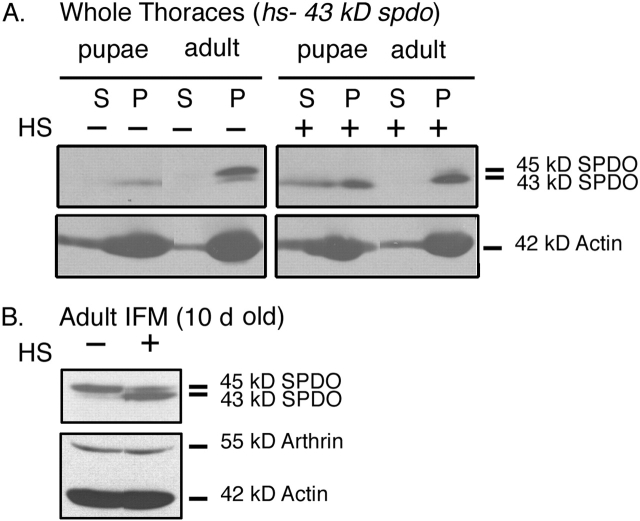
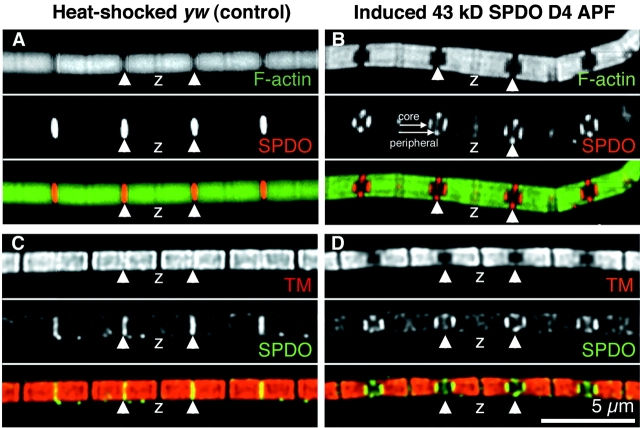
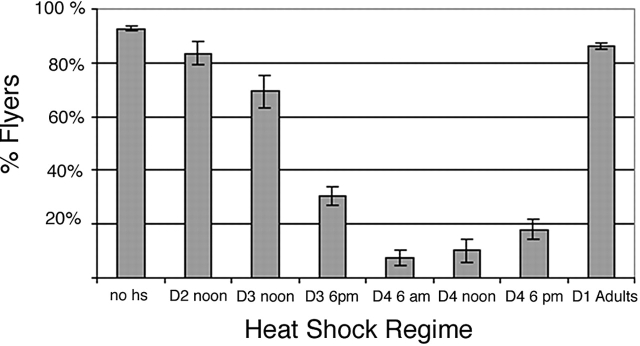
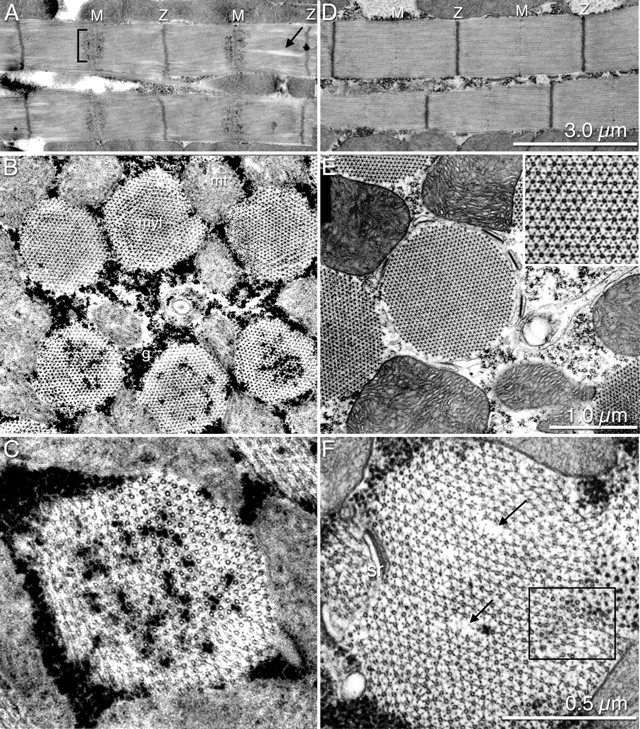
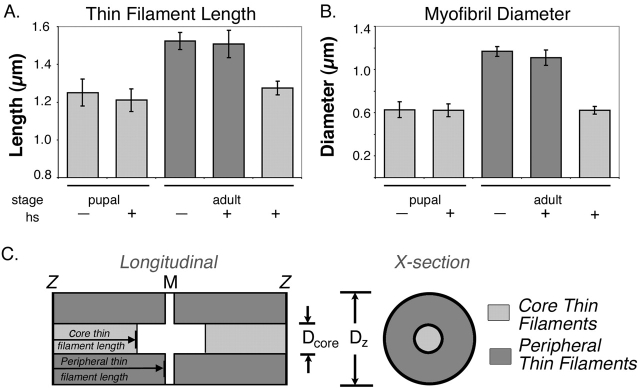
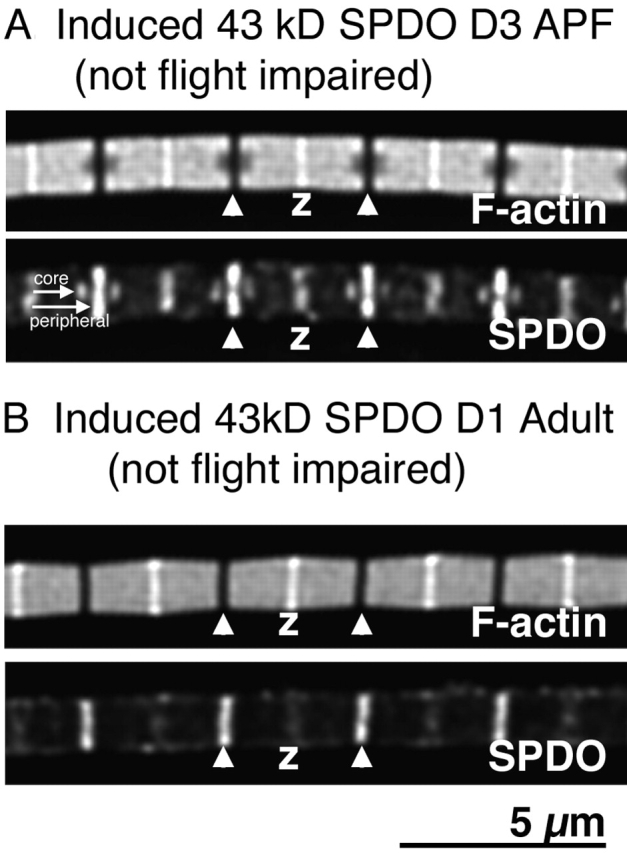
Tropomodulin (Tmod) is an actin pointed-end capping protein that regulates actin dynamics at thin filament pointed ends in striated muscle. Although pointed-end capping by Tmod controls thin filament lengths in assembled myofibrils, its role in length specification during de novo myofibril assembly is not established. We used the Drosophila Tmod homologue, sanpodo (spdo), to investigate Tmod's function during muscle development in the indirect flight muscle. SPDO was associated with the pointed ends of elongating thin filaments throughout myofibril assembly. Transient overexpression of SPDO during myofibril assembly irreversibly arrested elongation of preexisting thin filaments. However, the lengths of thin filaments assembled after SPDO levels had declined were normal. Flies with a preponderance of abnormally short thin filaments were unable to fly. We conclude that: (a) thin filaments elongate from their pointed ends during myofibril assembly; (b) pointed ends are dynamically capped at endogenous levels of SPDO so as to allow elongation; (c) a transient increase in SPDO levels during myofibril assembly converts SPDO from a dynamic to a permanent cap; and (d) developmental regulation of pointed-end capping during myofibril assembly is crucial for specification of final thin filament lengths, myofibril structure, and muscle function.
Figures
Similar articles
-
Mechanisms of thin filament assembly in embryonic chick cardiac myocytes: tropomodulin requires tropomyosin for assembly.J Cell Biol. 1995 May;129(3):683-95. doi: 10.1083/jcb.129.3.683. J Cell Biol. 1995. PMID: 7730404 Free PMC article.
-
Tropomodulin assembles early in myofibrillogenesis in chick skeletal muscle: evidence that thin filaments rearrange to form striated myofibrils.J Cell Sci. 1999 Apr;112 ( Pt 8):1111-23. doi: 10.1242/jcs.112.8.1111. J Cell Sci. 1999. PMID: 10085247
-
Actin dynamics at pointed ends regulates thin filament length in striated muscle.Nat Cell Biol. 2001 Jun;3(6):544-51. doi: 10.1038/35078517. Nat Cell Biol. 2001. PMID: 11389438
-
Capping actin filament growth: tropomodulin in muscle and nonmuscle cells.Soc Gen Physiol Ser. 1997;52:79-89. Soc Gen Physiol Ser. 1997. PMID: 9210222 Review.
-
Tropomodulin capping of actin filaments in striated muscle development and physiology.J Biomed Biotechnol. 2011;2011:103069. doi: 10.1155/2011/103069. Epub 2011 Oct 17. J Biomed Biotechnol. 2011. PMID: 22013379 Free PMC article. Review.
Cited by
-
The initial steps of myofibril assembly: integrins pave the way.Nat Rev Mol Cell Biol. 2009 Apr;10(4):293-8. doi: 10.1038/nrm2634. Epub 2009 Feb 4. Nat Rev Mol Cell Biol. 2009. PMID: 19190670 Review.
-
Deletion of Drosophila muscle LIM protein decreases flight muscle stiffness and power generation.Am J Physiol Cell Physiol. 2011 Aug;301(2):C373-82. doi: 10.1152/ajpcell.00206.2010. Epub 2011 May 11. Am J Physiol Cell Physiol. 2011. PMID: 21562304 Free PMC article.
-
Role of Actin-Binding Proteins in Skeletal Myogenesis.Cells. 2023 Oct 25;12(21):2523. doi: 10.3390/cells12212523. Cells. 2023. PMID: 37947600 Free PMC article. Review.
-
Drosophila Tropomodulin is required for multiple actin-dependent processes within developing myofibers.Development. 2023 Mar 15;150(6):dev201194. doi: 10.1242/dev.201194. Epub 2023 Mar 24. Development. 2023. PMID: 36806912 Free PMC article.
-
An ongoing role for structural sarcomeric components in maintaining Drosophila melanogaster muscle function and structure.PLoS One. 2014 Jun 10;9(6):e99362. doi: 10.1371/journal.pone.0099362. eCollection 2014. PLoS One. 2014. PMID: 24915196 Free PMC article.
References
-
- Almenar-Queralt, A., C.C. Gregorio, and V.M. Fowler. 1999. a. Tropomodulin assembles early in myofibrillogenesis in chick skeletal muscle: Evidence that thin filaments rearrange to form striated myofibrils. J. Cell Sci. 112:1112–1123. - PubMed
-
- Almenar-Queralt, A., A. Lee, C.A. Conley, L. Ribas de Pouplana, and V.M. Fowler. 1999. b. Identification of a novel tropomodulin isoform, skeletal tropomodulin, that caps actin filament pointed ends in fast skeletal muscle. J. Biol Chem. 274:28466–28475 (erratum published 275[17]:13164). - PubMed
-
- Ashburner, M. 1989. Drosophila, a Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Ball, E., C.C. Karlik, C.J. Beall, D.L. Saville, J.C. Sparrow, B. Bullard, and E.A. Fyrberg. 1987. Arthrin, a myofibrillar protein of insect flight muscle, is an actin-ubiquitin conjugate. Cell. 51:221–228. - PubMed
-
- Beall, C.J., M.A. Sepanski, and E.A. Fyrberg. 1989. Genetic dissection of Drosophila myofibril formation: effects of actin and myosin heavy chain null alleles. Genes Dev. 3:131–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
