

Multiple eIF4GI-specific protease activities present in uninfected and poliovirus-infected cells
- PMID: 11739682
- PMCID: PMC135685
- DOI: 10.1128/jvi.76.1.165-177.2002
Multiple eIF4GI-specific protease activities present in uninfected and poliovirus-infected cells
Abstract
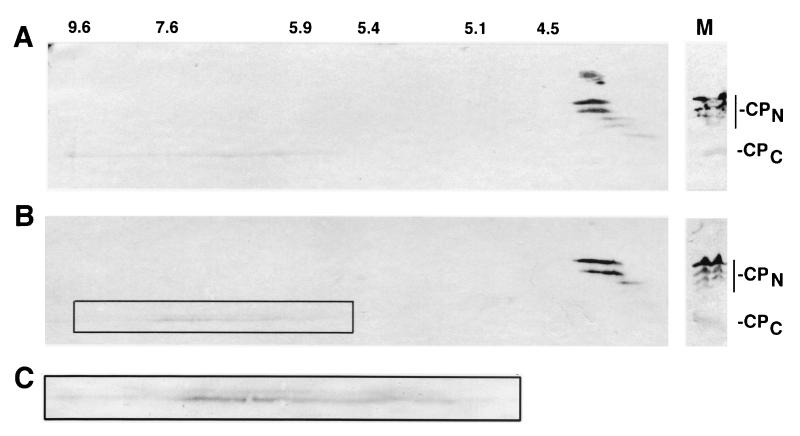
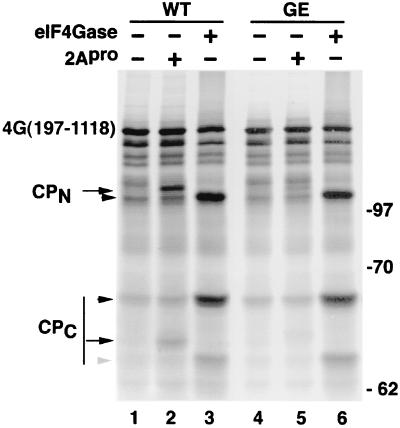
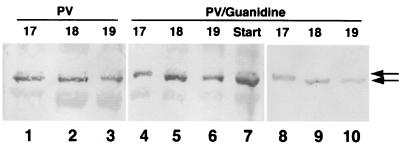
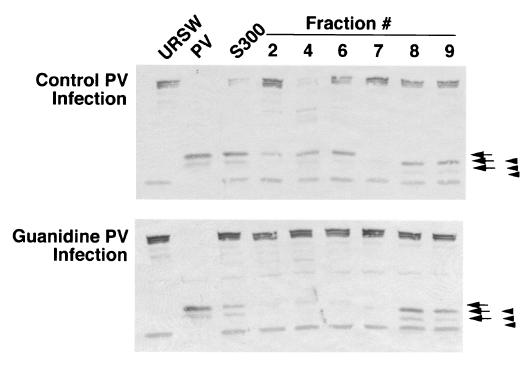
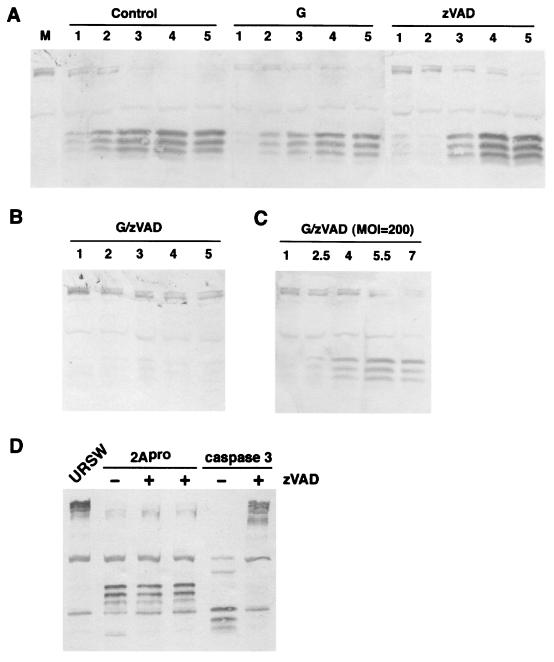
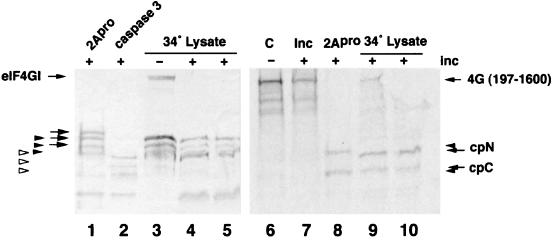
Cleavage of eukaryotic translation initiation factor 4GI (eIF4GI) is required for shutoff of host cell translation during poliovirus (PV) infection of HeLa cells. Reports published by several groups have led to confusion whether this cleavage is mediated by viral 2A protease (2A(pro)) or a putative cellular enzyme (termed eIF4Gase) which is activated by 2A(pro) or other aspects of viral infection. Here we have further investigated eIF4Gase activities in PV-infected cells. Column purification of eIF4GI cleavage activity separated two activities which generated N-terminal cleavage products of different lengths. Both activities were detected using either native eIF4G or radiolabeled recombinant eIF4G as the substrate. Analysis of cleavage products formed by each activity on native and mutant substrates suggests that one activity cleaves eIF4G1 at or very near the 2A(pro) cleavage site and the other activity cleaves approximately 40 residues upstream of the 2A(pro) cleavage site. When PV infections in HeLa cells were supplemented with 2 mM guanidine, which indirectly limits expression of 2A(pro), two distinct C-terminal cleavage fragments of eIF4GI were detected. These C-terminal cleavage fragments of eIF4GI were purified from infected cells, and a new eIF4GI cleavage site was mapped to a unique site 43 amino acids upstream of the known 2A(pro) cleavage site. Further, eIF4GI cleavage in vivo could be blocked by addition of zVAD to PV-guanidine infections. zVAD is a broad-spectrum caspase inhibitor which had no effect on 2A(pro) cleavage activity or PV polyprotein processing. Lastly, similar types of eIF4Gase cleavage activities were also detected in uninfected cells under various conditions, including early apoptosis or during cell cycle transit. The data suggest that the same types of eIF4GI cleavage activities which are generated in PV-infected cells can also be generated in the absence of virus. Taken together, the data support a model in which multiple cellular activities process eIF4GI in PV-infected cells, in addition to 2A(pro).
Figures




References
-
- Belsham, G., and R. Jackson. 2000. Translation initiation on picornavirus RNA, p. 869–900. In N. Sonenberg, J. Hershey, and M. Mathews (ed.), Translational control of gene expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
-
- Benton, P. A., J. W. Murphy, and R. E. Lloyd. 1995. K562 cell strains differ in their response to poliovirus infection. Virology 213:7–18. - PubMed
-
- Bovee, M. L., B. Lamphear, R. E. Rhoads, and R. E. Lloyd. 1998. Direct cleavage of eIF4G by poliovirus 2A protease is inefficient in vitro. Virology 245:241–249. - PubMed
-
- Bovee, M. L., W. E. Marissen, M. Zamora, and R. E. Lloyd. 1998. The predominant eIF4G-specific cleavage activity in poliovirus-infected HeLa cells is distinct from 2A protease. Virology 245:229–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
