

Cyclin T1 expression is mediated by a complex and constitutively active promoter and does not limit human immunodeficiency virus type 1 Tat function in unstimulated primary lymphocytes
- PMID: 11739686
- PMCID: PMC135689
- DOI: 10.1128/jvi.76.1.208-219.2002
Cyclin T1 expression is mediated by a complex and constitutively active promoter and does not limit human immunodeficiency virus type 1 Tat function in unstimulated primary lymphocytes
Abstract
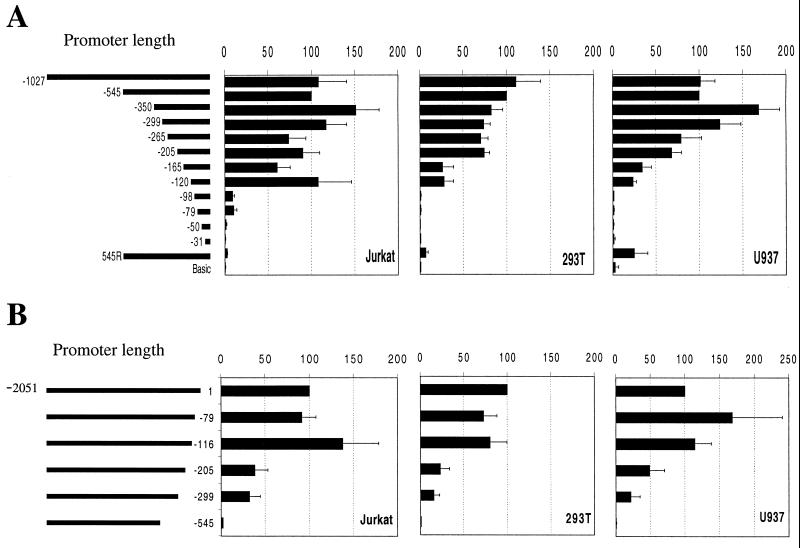
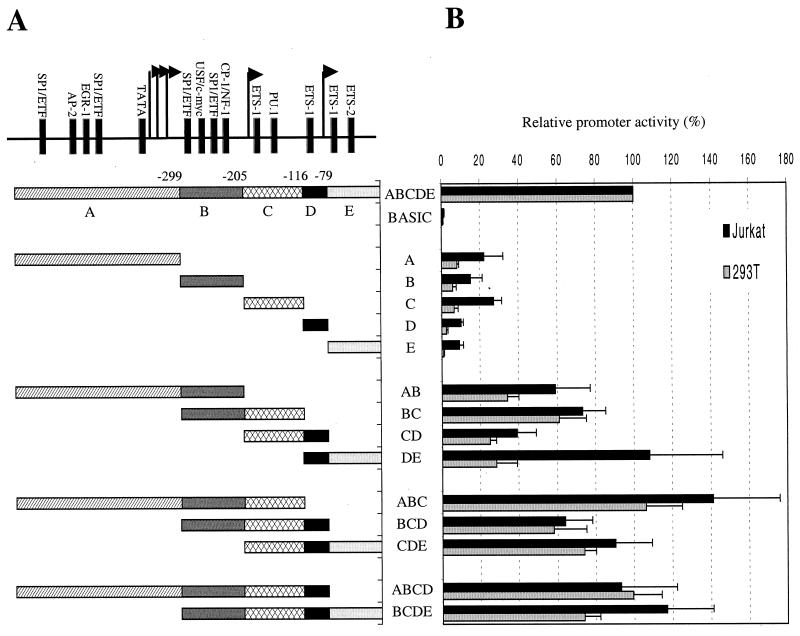
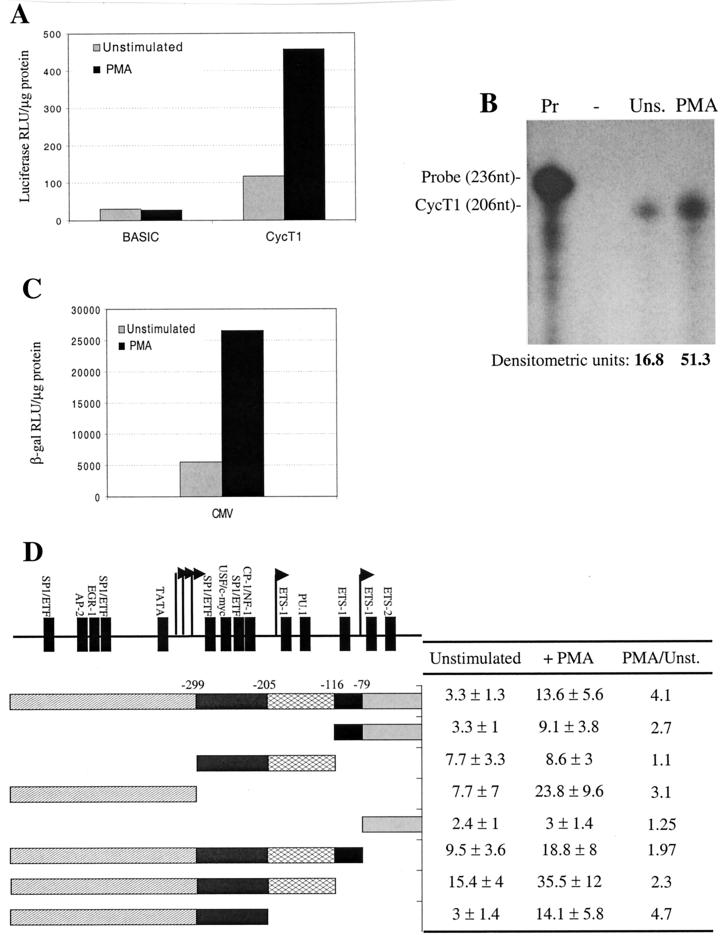
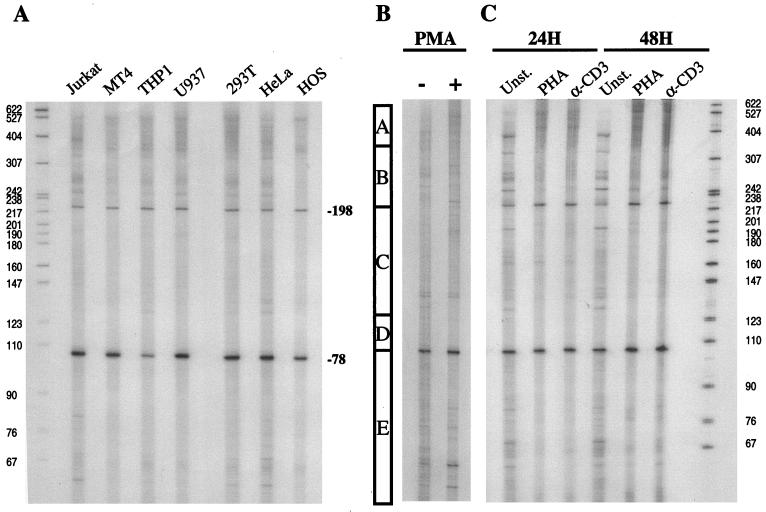
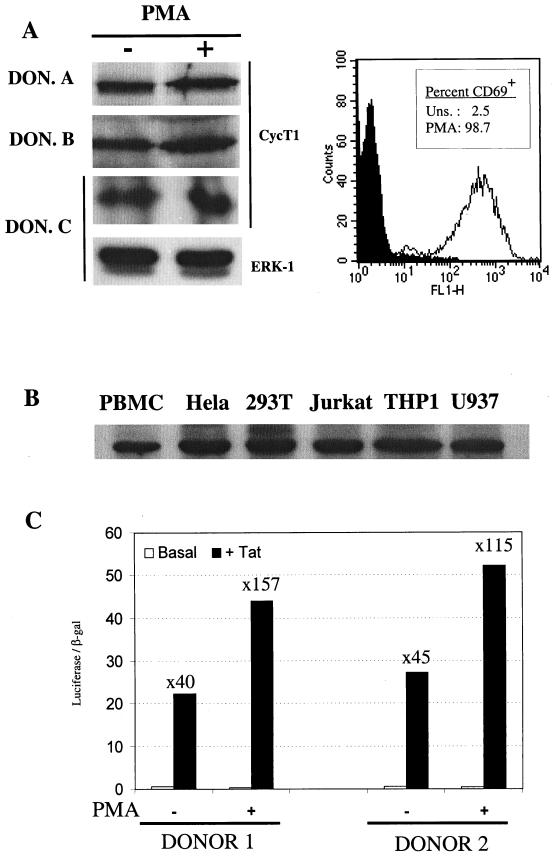
Cyclin T1 (CycT1), a component of positive-transcription-elongation factor-b (P-TEFb), is an essential cofactor for transcriptional activation by lentivirus Tat proteins. It is thought that low CycT1 expression levels restrict human immunodeficiency virus type 1 (HIV-1) expression levels and replication in resting CD4+ lymphocytes. In this study, we undertook a functional analysis of the cycT1 promoter to determine which, if any, promoter elements might be responsible for cellular activation state-dependent CycT1 expression. The cycT1 gene contains a complex promoter that exhibits an extreme degree of functional redundancy: five nonoverlapping fragments were found to exhibit significant promoter activity in immortalized cell lines, and these elements could interact in a synergistic or redundant manner to mediate cycT1 transcription. Reporter gene expression, mediated by the cycT1 promoter, was detectable in unstimulated transfected primary lymphocytes and multiple sites within the promoter could serve to initiate transcription. While utilization of these start sites was significantly altered by the application of exogenous stimuli to primary lymphocytes and two distinct promoter elements exhibited enhanced activity in the presence of phorbol ester, overall cycT1 transcription was only modestly enhanced in response to cell activation. These observations prompted a reexamination of CycT1 protein expression in primary lymphocytes. In fact, steady-state CycT1 expression is only slightly lower in unstimulated lymphocytes compared to phorbol ester-treated cells or a panel of immortalized cell lines. Importantly, CycT1 is expressed at sufficient levels in unstimulated primary cells to support robust Tat activity. These results strongly suggest that CycT1 expression levels in unstimulated primary lymphocytes do not profoundly limit HIV-1 gene expression or provide an adequate mechanistic explanation for proviral latency in vivo.
Figures

References
-
- Adams, M., L. Sharmeen, J. Kimpton, J. M. Romeo, J. V. Garcia, B. M. Peterlin, M. Groudine, and M. Emerman. 1994. Cellular latency in human immunodeficiency virus-infected individuals with high CD4 levels can be detected by the presence of promoter-proximal transcripts. Proc. Natl. Acad. Sci. USA 91:3862–3866. - PMC - PubMed
-
- Albrecht, T. R., L. H. Lund, and M. A. Garcia-Blanco. 2000. Canine cyclin T1 rescues equine infectious anemia virus tat trans-activation in human cells. Virology 268:7–11. - PubMed
-
- Alcami, J., T. Lain de Lera, L. Folgueira, M. A. Pedraza, J. M. Jacque, F. Bachelerie, A. R. Noriega, R. T. Hay, D. Harrich, R. B. Gaynor, et al. 1995. Absolute dependence on kappa B responsive elements for initiation and Tat-mediated amplification of HIV transcription in blood CD4 T lymphocytes. EMBO J. 14:1552–1560. - PMC - PubMed
-
- Antequera, F., and A. Bird. 1999. CpG islands as genomic footprints of promoters that are associated with replication origins. Curr. Biol. 9:R661–R667. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
